| English | Главная | Сотрудники | Экспедиции | Ссылки | Публикации | События | Потери науки |
ПРОЦЕСС ЭВОЛЮЦИИ И МЕТОДОЛОГИЯ СИСТЕМАТИКИ
А.П. Расницын
ОТ АВТОРА
Эта книга представляет некоторый итог моих размышлений о теоретических основах таксономического круга биологических дисциплин. В этот круг я включаю разделы биологии, анализирующие организменный уровень структуры и происхождения биологического разнообразия. Структуру биоразнообразия на этом уровне разрабатывает систематика с ее методологическим и лингвистическим аппаратами (таксономией и таксономической номенклатурой), происхождение биоразнообразия - эволюционная теория и филогенетика (методология конкретных филогенетических исследований).
Интерес к эволюционной теории, наверное, не требует объяснения: в начале моего научного пути (1950е-1960е годы) им было заражено едва ли не большинство биологов. Что до методологии, обратиться к ней меня вынудили два разных обстоятельства. Одно из них - давний кризис таксономии, выражающийся в нескончаемом конфликте между "традиционной" систематикой и соперничающими подходами - фенетикой и, позднее, кладизмом. Интуитивно чувствуя преимущество традиционной систематики, я тоже включился в эту дискуссию. Результаты моих попыток развить аргументацию этих споров представлены ниже в разделах, посвященных филогенетике и систематике.
В случае с номенклатурой обстоятельства были иными и коренились в моей палеонтологической деятельности. Хотя систематика и номенклатура имеют долгую историю, базовые принципы номенклатуры на мой взгляд еще далеко не все выявлены и сформулированы. Некоторые из них настолько просты и самоочевидны, что их существование и действие ощутимо лишь в периферических областях систематики. Когда мы вынуждены классифицировать трудные объекты, лишенные многих или всех таксономически важных признаков, мы часто вынуждены нарушать правила номенклатуры. Это обычно в палеонтологии, в изучении паразитических червей и бесполых форм грибов, где исследователь не может избежать нарушения нормальной таксономической практики и соответствующих правил номенклатуры. Такая деятельность не приносит удовлетворения, пока исследователь не начинает различать более глубокие основания того, почему он не может следовать общепринятой практике. Причина лежит в несоответствии стандартных правил номенклатуры таксономически неполноценному материалу, который тем не менее подлежит таксономической обработке. Это противоречие делает базовые принципы номенклатуры лучше видимыми и потому доступными выявлению и изучению (Расницын, 1986а, 1992b).
До недавнего времени мои рассуждения на эти две главные темы, т.е. об онтологии эволюционного процесса (Расницын, 1971а,б, 1972а, 1975, 1984, 1986б, 1987, 1988а,б, 1989а-г) и методологии анализа его результатов (Пoнoмapeнкo, Pacницын, 1971, Расницын, 1972б, 1983а, 1986а, 1988в, 1990а, 1992а,б, Rasnitsyn, 1982, 1987, 1991, 1996a, 2000, Pacницын, Длyccкий, 1988, Емельянов, Pacницын, 1991) текли изолированно, и лишь недавно я понял, как тесно одно зависит от другого (Pacницын, 2002). По счастливому совпадению именно в это время я получил любезное предложение от Российского Энтомологического Общества изложить свою точку зрения на методологию филогенетического и таксономического исследования в одном из выпусков трудов Общества, каковое с удовольствием и выполняю. Пользуюсь случаем выразить свою искреннюю признательность многочисленным коллегам из Палеонтологического и Зоологического институтов РАН, Московского и Санкт-Петербургского университетов и многих других коллег в России и за рубежом, коих слишком много, чтобы перечислять, за стимуляцию к размышлениям, обсуждение и критику результатов этих размышлений. Особо хочу отметить А.Г. Пономаренко, С.П. Расницына, А.С. Раутиана и покойного В.В. Жерихина, дискуссии с которыми было особенно бурными и плодотворными, и Г.С. Медведева за предоставленную привилегию опубликовать эту книгу в Трудах РЭО.
ОНТОЛОГИЯ ЭВОЛЮЦИИ: ДВЕ ПАРАДИГМЫ
В современной теории эволюции конкурируют два базовых подхода, две парадигмы - редукционистская, сводящая эффекты на уровне целого к явлениям на уровне его элементов, и антиредукционистская, анализирующая процессы низших уровней, исходя из свойств целого. Я здесь пользуюсь привычной терминологией, но она не точна - хотя бы потому, что в действительности оба подхода редукционистские. Мы не в состоянии оперировать явлениями реальности во всей неисчерпаемости их содержания и поэтому всегда вынуждены редуцировать это богатство тем или иным способом. Разница лишь в том, что один подход редуцирует систему вниз, к элементам, другой вверх, к целому (Мейен, 2001: 382-383). Однако такого термина, как "элементаризм" или что-то подобное, кажется, не предложено, а вводить его здесь мне кажется неуместным. Поэтому попробую обойтись упомянутыми старыми названиями. "Редукционистскую" парадигму представляет синтетическая теория эволюции, восходящая к А. Вейсману и Т. Моргану, "антиредукционистскую" парадигму - эпигенетическая теория, возникшая усилиями Р. Гольдшмита, А.Г. Гурвича, К.Х. Уодингтона и И.И. Шмальгаузена. Взаимоотношения этих двух направлений специально рассмотрены М.А. Шишкиным (1987, 1988а) и Д.Л. Гродницким (2000, 2002). Опираясь на их результаты, попытаюсь изложить свое видение этой проблемы.
СИНТЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ
Синтетическая, а более точно - популяционно-генетическая теория эволюции (ибо синтез там скорее прокламирован, чем достигнут, см. Шишкин, 1987, 1988а) рассматривает наследственность, т.е. способность к устойчивому воспроизводству фенотипа и его признаков в последовательных поколениях, как свойство особых элементарных носителей-генов, ныне отождествляемых с участками (локусами) хромосом и далее с участками ДНК (цистроны и т.п.). В соответствии с этим эволюционный процесс представляется как динамика аллельных частот в популяциях, контролируемая отбором (через оценку жизнеспособности и, соответственно, дифференциальное воспроизводство фенотипов) и стохастическими процессами. Признаки возникают и изменяются в результате мутаций и рекомбинаций генов. Поскольку все эти изменения не обладают свойством изначальной целесообразности, именно отбор формирует состав популяций и свойства организмов, и делает он это с точностью, ограниченной лишь стохастическими факторами. Кроме этого фактора случайности, единственное препятствие такому соответствию - обмен генами, который унифицирует структуру популяций на уровне, отвечающем не локальным, а усредненным характеристикам отбора.
В соответствии с этими взглядами организация живых существ представляется мягкой глиной в руках отбора или, точнее, набором признаков, свободно тасуемых отбором, а эволюционный процесс на надвидовом уровне - равномерным в той мере, в какой равномерны определяемые средой вариации отбора. Эта равномерность нарушается лишь дивергенциями и вымираниями, каковые только и могут быть использованы для построения естественной системы организмов.
Синтетическая теория, конечно, признает и плейотропность генов (влияние каждого из них на многие признаки) и полигенность признаков (множественную генетическую обусловленность каждого из них), так что каждый признак в действительности зависит от всего генома (например, в гипотезе генного баланса). Однако эти признания в действительности декларативны, так как тотальная взаимозависимость элементов структуры и функций организма и, соответственно, отсутствие сколько-нибудь постоянного вклада гена в приспособленность делают невозможной редукцию эволюционного процесса к уровню элементарных генетических (тем более молекулярно-генетических) событий и их частот.
Все эти упрощения ведут к значительному несоответствию между предсказаниями синтетической теории и результатами наблюдений. Приведем лишь немногие из таких несоответствий, касающихся надорганизменного (в основном таксономического) уровня анализа. Проблемы, связанные непосредственно с онтогенезом, его механизмами и наследственным обеспечением, подробно рассмотрены Шишкиным (1987, 1988а,б) и Раутианом (1993).
Обратимся прежде всего к темпам эволюционного процесса, оцениваемым по скорости возникновения и вымирания таксонов того или иного ранга. Конечно, этот метод оценки несовершенен: хотя таксон - единственное понятие, самым смыслом существования которого является попытка интегрированной оценки сходств и различий организмов, сопоставимость разных одноранговых таксонов не обеспечена ничем, кроме интуиции систематиков (см. ниже). Однако других способов оценки темпов эволюции организмов у нас нет: сколь ни несовершенна (субъективна) оценка различий в таксономических рангах, она более информативна, чем более объективная оценка различий с помощью признаков. Хорошо известно, что внешне одинаковые различия могут достигаться настолько разными способами и оказываются проявлениями насколько разномасштабных преобразований организации, что оценка темпов эволюции с их помощью будет только вводить в заблуждение. Особенно ярко подтверждают этот вывод материалы по эволюции в островных условиях и на ранних этапах эволюции таксонов (феномен архаического многообразия), когда быстро и легко возникают гротескные формы с несбалансированной организацией, чьи признаки в норме (в стабилизированных группах) характеризуют таксоны весьма высокого ранга (подробнее см. ниже). В то же время палеонтологические данные относительно темпов таксономической эволюции в разных группах организмов дают результаты и обнаруживает тенденции, которые вряд ли можно в сколько-нибудь заметной степени списать на несопоставимость одноранговых таксонов (Расницын. 1987).
Согласно синтетической теории эволюционный процесс управляется в основном теми же факторами и механизмами, которыми оперирует генетика популяций. В этом случае и скорость макроэволюции должна подчиняться тем же закономерностям, что скорость изменения частот генов в генетике популяций, т. е. определяться мощностью потока адаптивно компетентных генетических вариаций, протекающих через эволюционирующую совокупность ("Основное уравнение эволюции: скорость эволюции в пределах некоей популяции равна числу мутаций, возникающих в единицу времени, умноженному на долю фиксируемых мутаций"; Вилсон, 1985). Следовательно, скорость должна быть тем выше, чем выше частота мутаций и скорость смены поколений, чем больше размер популяций и мобилизационный резерв изменчивости.
Палеонтологический материал не позволяет прямо оценить большинство перечисленных параметров, используемых в генетике популяций. Некоторые из них, коррелирующие с другими признаками (размеры, таксономическая принадлежность), все же могут быть косвенно оценены у ископаемых. В частности, если генетический подход к эволюции справедлив, то ее скорость у млекопитающих, обладающих сравнительно медленной сменой поколений и небольшими популяциями (в связи с относительно крупными размерами особи), должна быть минимальной, особенно у наиболее крупных из них, у одноклеточных - максимальной, а у беспозвоночных - в среднем промежуточной. В действительности же, как показывают многочисленные палеонтологические данные, эти соотношения имеют обратный характер (Расницын, 1987). При этом различия весьма велики даже на уровне вида, таксономической категории, которая считается наиболее сравнимой в разных группах. Так, возраст (в миллионах лет) полусовременной фауны (в которой половина видов современные, половина - вымершие) у крупных млекопитающих (хоботные и копытные) 0,2, у мелких 0,5, у птиц и рыб 0,7, у насекомых 3-7, у моллюсков 3,5-5, у диатомовых водорослей - 15. Время полувымирания (по аналогии с полураспадом, время вымирания половины исходного числа видов) в тех же единицах наименьшее у слонов (0,18), а в среднем у млекопитающих 0,54, у костистых рыб 3,5, у граптолитов 1,3, иглокожих 4,2, двустворок 7, у планктонных фораминифер 5, бентосных 18-24, у диатомовых водорослей 5,5, у динофлагеллат 9. На старших таксономических уровнях различия подобные, но часто более резкие. Например, возраст полусовременной фауны для родов млекопитающих 4 млн. лет, птиц 10, рептилий 20, рыб 30-50, насекомых 40, моллюсков 60, фораминифер 230.
Полученные цифры не могут поставить под сомнение обоснованность генетических предсказаний для своего уровня анализа. Отбор абсолютно автоматичен: если поток вариаций вызывает появление в популяции особей, различающихся по степени приспособленности, они будут неуклонно отбираться в полном соответствии с положениями генетики популяций. В той мере, в какой эти различия наследуемые, разное участие особей в воспроизводстве популяции столь же автоматически (с учетом стохастики) приведет к изменению наследственной структуры популяции во времени. Следовательно, предсказываемое генетическими теориями распределение скоростей эволюции обязательно должно иметь место в природе в той мере, в которой мощность общего потока генетических вариаций в долговременном аспекте коррелирована с мощностью потока отбираемых (повышенно адаптивных) наследуемых изменений, фиксация которых и составляет процесс изменения популяций. Наглядной иллюстрацией справедливости популяционно-генетических предсказаний могут служить хорошо известные различия по типичным скоростям селекции между породами домашних животных, сортами растений и штаммами микроорганизмов. Не менее показательны и различия между обычными (измеряемыми миллионами лет, см. выше) и потенциальными темпами эволюции насекомых, когда на острове Гавайи возрастом 0,5 млн. лет. (Rotondo et al., 1981) в результате 17 вселений возникло 46 видов жука усача рода Plagithmysus (Gressit, 1978), т.е. не менее одного акта видообразования происходило в среднем за 150-200 тыс. лет. Г.Х. Шапошников почти вывел новый вид тли всего за один сезон (см. ниже).
Столь же очевидно, однако, что распределение скоростей эволюции на уровне вида и выше в первом приближении противоположно предсказанному популяционной генетикой. Это означает, что популяционно-генетические ограничения не являются таковыми для эволюционного процесса. Грубо говоря, поток общей генетической изменчивости эволюционно избыточен даже в популяциях крупных млекопитающих, не говоря уже о других, более обильных особями и быстрее плодящихся организмах. Скорость изменений на надпопуляционном уровне (скорость эволюции) должна регулироваться какими-то другими факторами. Вопрос о том, какие это могут быть факторы, анализируется в рамках альтернативной парадигмы (см. ниже). Здесь же можно заключить, что тот уровень, свести к которому эволюционный процесс и тем самым его объяснить пытается синтетическая теория эволюции, не отвечает возлагаемым на него надеждам и не дает ожидаемых объяснений - по крайней мере в отношении парадокса эволюционных скоростей.
Существуют и другие группы данных, не укладывающиеся в рамки синтетической теории эволюции. Если организация живых существ - мягкая глина в руках отбора или набор признаков, свободно тасуемых отбором, то ее результаты должны быть предсказуемыми в той мере, в какой известны условия, в которых протекала эволюция. Этого не наблюдается, что успешно демонстрируют нам антиселекционисты на многочисленных примерах (см., например, Любищев, 1982). Даже близкородственные формы в сходных, казалось бы, условиях, могут вести себя совершенно по-разному. Самый парадоксальный пример - крайне экстравагантная в репродуктивной сфере пятнистая гиена и вполне ординарная в этом отношении, близкая к ней полосатая гиена.
Еще менее объяснима с синтетической точки зрения широко распространенная, если не универсальная дискретность биологического разнообразия. В синтетико-эволюционной парадигме естественна лишь одна форма дискретности - дискретность амфимиктического вида в условиях, допускающих обмен генами, достаточный для унификации состава включенных в обмен популяций вопреки локальным вариациям давления отбора. Отсюда следует, что между степенью (полнотой и древностью) изоляции и степенью дивергенции должна существовать хорошая корреляция. Этого в действительности не наблюдается: тормозящее (нивелирующее различия) влияние обмена генами на скорость дивергенции подтвердить не удается. К такому выводу пришел, в частности, С.С. Шварц (1980), сравнивавший виды млекопитающих с разной склонностью образовывать изоляты.
С точки зрения синтетической теории эволюции совершенно необъяснима дискретность видов у партеногенетических и бесполых организмов, лишенных обмена генами, но она имеет место. Сравнение близких обоеполых и партеногенетических видов у коловраток (Майр, 1974) и жуков-долгоносиков (Иванова, 1978; В.В. Жерихин, личное сообщение), обоеполых и бесполых видов у протистов (Ю. Полянский, 1957; Poljansky, 1977), низших водорослей (В. Полянский, 1956) и папоротников (Farrar, 1990) показало, что однополые и бесполые виды столь же дискретны, как и обоеполые.
Не менее показателен длительный (миллионы, десятки и возможно даже сотни миллионов лет) эволюционный стазис на видовом и родовом уровне, когда об обмене наследственной информацией не может быть и речи. Так, популяции четырех или пяти из тридцати австралийских видов бессяжковых насекомых (Protura) на видовом уровне неотличимы от популяций с далеких материков и островов - Калимантан, Япония, Южная Африка, Европа (Tuxen, 1967). Бессяжковые нестойки к высыханию и не покидают почву, поэтому трансконтинентальные миграции для них практически исключены, и, объясняя их распространение, нам, очевидно, не избежать ссылок на дрейф континентов (тем более что для бессяжковых известен и классический "дрейфовый" ареал вида, охватывающий оба побережья Атлантического океана, у Delamarentulus tristani Silv., распространенного на Коста-Рике и в Западной Африке; Tuxen, 1963). Но из этого автоматически следует возраст вида у бессяжковых, оцениваемый многими десятками миллионов лет.
Известны и более прямые указания на большую древность некоторых видов. Так, в фауне эоценового балтийского янтаря (возраст не менее 40 млн. лет) сейчас известно несколько видов насекомых и около десятка видов клещей, на видовом уровне неотличимых от современных; более обычно персистирование на интервалах в 10-20 млн. лет (Zherikhin, 1999). Находки разнообразных плиоценовых галлов, неотличимых от галлов, вызываемых современными видами насекомых на тех же растениях, гораздо моложе (3-5 млн. лет), но они не менее важны, так как подтверждают широкое распространение не только морфологического, но и биохимического стазиса насекомых в течение миллионов лет (Zherikhin, 2002a). Многие из этих находок происходят из Северной Европы, так что речь идет о видах, переживших ледниковый период, но не изменившихся даже биохимически (поскольку морфологическая специфика галла обусловлена биохимическим воздействием насекомого-галлообразователя на ткани растения-хозяина). Таким образом, стазис длиной в миллионы лет осуществляется не только в отсутствие обмена генами, но и вопреки несомненным глубоким, возможно даже катастрофическим, изменениям условий.
Рекордсменом, однако, является щитень Triops cancriformis (Schaffer), который практически не изменился с раннего триаса (около 230 млн. лет), и даже вопрос о видовой самостоятельности пермских популяций остается нерешенным (Tasch, 1969). В подтверждение этого вывода данные молекулярных часов для двух морфологически едва различимых японских популяций другого вида, T. longicaudatus (LeConte) свидетельствуют о их расхождении около 15 млн. лет назад. Для видов этого рода, включая T. cancriformis, дивергенции датируются 25-45 млн. лет (Suno-Uchi et al., 1997; в расчетах использовались часы, откалиброванные специально для ракообразных).
Еще менее понятен в этой системе взглядов часто наблюдающийся феномен, когда старшие таксоны оказываются более дискретными и демонстрируют более ясные родственные отношения, чем виды. Действительно, виды расходились относительно недавно, имели меньше времени на дивергенцию и, следовательно, должны были сохранить больше следов того, как именно протекала дивергенция (подробнее см. ниже).
Все сказанное, на мой взгляд, достаточно убедительно показывает серьезное несоответствие между популяционно-генетическим подходом и его следствиями - синтетической теорией эволюции и биологической концепцией вида, с одной стороны, и совокупностью наблюдений относительно структуры и эволюции биологического разнообразия, с другой. Поэтому пора уже перейти к альтернативной системе взглядов, к другой парадигме.
ЭПИГЕНЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ
В отличие от синтетической, эпигенетическая теория видит эволюционный процесс прежде всего как процесс эволюционного преобразования онтогенеза (Шишкин, 1987, 1988 а,б; Раутиан, 1993). При этом особое внимание обращается на целостность онтогенеза в том смысле, что и сам процесс, и его результат (строение организма на последовательных этапах его развития) гораздо устойчивее, чем любые отдельные факторы и процессы развития. Нормальное развитие эквифинально и способно релаксировать (подавлять, поглощать, нивелировать) очень широкий круг воздействий и возмущений, как внешних, так и внутренних (включая результаты мутаций и рекомбинаций, равно как и иных ошибок и нарушений нормального развития).
Таким образом, наследуется (устойчиво воспроизводится в последующих поколениях) скорее нормальный онтогенез (норма реакции) в целом, чем отдельные признаки. Помимо нормы, существует масса разнообразных уклонений (аберраций) развития с неустойчивым воспроизведением, которые в нормальных условиях реализуются редко. В неблагоприятных условиях, когда в результате ли сильного внешнего воздействия или по внутренним причинам (ошибки развития, включая мутации, рекомбинации, гибридизацию, создающие несбалансированный эпигенотип), механизмы защиты нормы удается нарушить или преодолеть, развитие идет по аберрантному (уклоняющемуся и неустойчивому) пути. Поскольку эндогенные аберрации развития легко возникают не только при половом размножении, но и при партеногенетическом и бесполом, их возникновение приходится интерпретировать как результат вскрытия скрытой гетерогенности популяции (сохраняющейся и при клонировании), а не возникновение de novo каких-то генетических изменений.
Именно аберрации рассматриваются эпигенетической теорией как материал отбора, который способен либо подавить ее реализацию более эффективно, чем прежде, либо создать на ее основе новую адаптивную норму. Этот процесс легче описать в рамках метафоры эпигенетического ландшафта. Нормальный онтогенез представляет собой устойчивую (стабилизированную) последовательность тесно связанных отдельных эпигенетических процессов. Более или менее обособленные отрезки этой последовательности именуют креодами. Если представить себе креод в виде сильно заглубленной долины в некотором ландшафте, по уклону которой течет онтогенетический процесс (Рис. 1), то аберрациями будут пологие боковые долинки, приподнятые в бортах главной. Чтобы аберрация реализовалась, т.е. чтобы онтогенез мог выйти в боковую долину, нужно либо сильное внешнее воздействие на развивающийся организм, выталкивающее его на боковую долину (Рис. 2а), либо изменение самого ландшафта, выполаживающее дно долины (например, в результате крупной мутации, Рис. 2б), либо то и другое вместе (Рис. 2в).

Рис. 1. Участок эпигенетического ландшафта, показывающий перепады высоты стенок (порогов устойчивости) креода в местах ответвления аберративных долин. А1, А2 - аберративные долины, N - главная долина (креод) (по Шишкину, 1987).

Рис. 2. Соотношения между строением эпигенетического ландшафта и характером повреждающего воздействия: а - уклонение развития на боковую долину за счет сильного внешнего воздействия (длинная стрелка); б- такое же уклонение под действием сильной мутации, вызывающей нарушение креода; в - промежуточное состояние (по Шишкину, 1987).
Аберрации мало устойчивы ("не наследственны"; более распространенные из них часто именуются модификациями), но они постоянно, хотя и непредсказуемо в деталях, воспроизводятся в популяции, поскольку эпигенетический ландшафт с соответствующими боковыми долинками есть адаптивная норма вида. Поэтому, несмотря на свою слабую наследуемость, именно аберрации служат материалом отбора: если аберрация оказалась полезной, отбор будет избирательно сохранять те эпигенотипы, которые более устойчиво ее воспроизводят. Другими словами, преимущество получат те эпигенотипы, где соответствующая долинка углублена, и уровень ее дна на стыке с главной долиной более приближен ко дну последней. Если при этом еще и главная долина будет выполаживаться ниже развилка (т.е. если понизится устойчивость прежней нормы), прежняя аберрация будет иметь шанс стать новой нормой. Если же прежняя норма сохранит свое значение, то стабилизируются обе нормы и будет выработан механизм онтогенетического переключения между ними. Если этот механизм использует для переключения средовой сигнал, мы получаем типичную модификационную изменчивость, если же сигнал будет генетическим (например, рекомбинация), мы получим одну из форм менделирования. Таким путем менделирующие признаки возникают и в природе (например, обычные механизмы определения пола), и в лаборатории (например, в процессе стабилизации чистых линий).
В целом для эпигенетической теории эволюции зафиксированный ("запомненный") в адаптивной норме спектр возможных путей развития (креодов и их аберраций), т.е. эпигенетический ландшафт - это сущность живого организма, его онтогенетический потенциал, то, что определяет существование организма и с чем работает отбор. Чисто генетический уровень (гены и их аллели, динамика их частот в популяции, мутации, рекомбинация и т. п.) лежит гораздо глубже и не определяет специфики эволюционных процессов, так же как из процессов и закономерностей квантовомеханического уровня не вывести специфики происходящего на макроуровне. "Гены приходят и уходят, а креоды остаются": хорошо известно, что признаки более устойчивы, чем их гены и аллели, их будто бы определяющие.
Более подробно онтогенетические процессы в аспекте наследования рассмотрены М.А. Шишкиным (1987, 1988а). Для нас же более важна другая сторона проблемы эволюции онтогенеза.
Адаптивный компромисс
Целостность организации живых существ оказывает существеннейшее влияние на характер их эволюции. На морфогенетическом уровне это влияние проявляется в том, что из-за глубокой взаимозависимости между изменениями разных креодов и, как следствие, между изменениями разных свойств и признаков организма креоды оказываются высоко стабилизированными. Они с большим трудом поддаются изменению за пределы нормальной внутривидовой изменчивость, которая сама уже стабилизирована отбором предыдущих поколений. Поэтому эволюция, т.е. успешный выход за пределы нормального, стабилизированного эпигенотипа, происходит с трудом, нечасто и с мало предсказуемым результатом.
Другими словами, в руках отбора организация живых существ оказывается хрупким, капризным материалом, эволюционные преобразования - более или менее скачкообразными, а возникающее в результате биоразнообразие - более или менее дискретным. Эта дискретность не абсолютна, и даже там, где она существует, нам не всегда удается ее легко нащупать. Но она существует и очень широко распространена, чего биологи и особенно систематики не слишком склонны подчеркивать, сосредоточившись на исключениях и трудных случаях. Существование же исключений вполне естественно. Стабилизированный эпигенотип (адаптивная норма) включает в себя определенный, более или менее широкий спектр изменчивости, такой же результат отбора предшествующих поколений, как и модальные характеристики фенотипа. Спектры нормальной изменчивости близких видов вполне могут перекрываться, создавая проблемы для систематиков, даже если сами эпигенотипы при этом различаются дискретно.
Проблема в том, что мы пока не знаем, как увидеть, очертить и измерить сам по себе эпигенотип, а не только его внешние проявления. Подобно атому или нейтрино, он недоступен прямому наблюдению, но он почти так же надежно выводится из имеющихся данных. А недоступен он прямому наблюдению не из-за малости размеров, а из-за того, что эпигенотип - это очень сложная система взаимосвязей и взаимодействий. Но поскольку эпигенотип, подобно атомному заряду, существенно предопределяет свойства и поведение его воплощений (особей, видов или иных таксонов), его можно определить еще и как материализованную сущность всех этих воплощений. Так что эпигенетическая теория эволюции - это в некотором смысле возвращение не только к типологии, столь яростно обруганной сторонниками биологической концепции вида, но и к эссенциализму - философскому направлению, еще менее респектабельному, чем типология, из-за своих идеалистических корней. Однако мой эсенциализм существенно иного толка, чем у Платона, поскольку сущность здесь признается материальной и познаваемой.
Однако вернемся к эволюции эпигенотипа. Высокая связность эпигенотипа и забуференность его эволюционных изменений удобно анализировать в рамках еще одной метафоры, метафоры адаптивного компромисса (Расницын, 1987). Действительно, устойчивость сбалансированного эпигенотипа можно рассматривать как одно из следствий теории систем, в частности, того ее утверждения, что ни одна система не может быть оптимизирована одновременно более чем по одному параметру. Оптимизация реальных систем возможна лишь как нахождение компромисса между противоречивыми требованиями оптимизации различных параметров. Для живых существ с глубоко пронизывающими всю их организацию корреляциями и взаимозависимостями компромисс между различными адаптивными функциями должен быть особенно напряженным. Поэтому устойчивый эпигенотип должен быть организован по принципу глубоко проработанного компромисса между противоречивыми потребностями максимальной оптимизации всех адаптивных функций.
Отношение к живому существу как к адаптивному компромиссу не является в сущности чем-то новым. Э. Майр имел в виду, по-видимому, именно его, когда писал, что "эпигенотип вида, его система канализаций развития и обратных связей часто столь хорошо интегрирована, что с замечательным упорством противостоит изменениям" (1974, с. 353). Н.Н. Воронцов (1961, 1963), показав, что обеспечение адаптивной функции может совершенствоваться разными способами, обнаружил при этом, что максимально эффективные варианты различных подсистем почему-то лишь ограниченно совместимы и не встречаются все вместе в одном организме. Очевидно, потому, что найти компромисс между всеми ими пока не удалось. Значительно раньше близкие идеи борьбы (конкуренции) частей в организме высказывали В. Ру и А. Вейсман (Вейсман, 1905), до них в виде принципа компенсации или уравновешивания - И.В. Гете и Ж. Сент Илер (см.: Дарвин, 1991, с. 128), а еще раньше как принцип экономии - Аристотель (1937).
Метафора адаптивного компромисса важна тем, что из нее можно вывести широкий круг сравнительно легко проверяемых следствий. Наиболее общее из них - это трудность изменения хорошо сбалансированной организации. Конечно, эта трудность не абсолютна, и в той мере, в какой изменения осуществимы, утрата адаптивного значения какой-либо функции будет вести к редукции обеспечивающих ее систем. Утрата приспособления, потерявшего свое значение, естественно объяснить тем, что редукция или дезинтеграция ставших ненужными систем позволяет дополнительно оптимизировать другие системы, сохранившие адаптивность. Это объяснение представляется более естественным, чем привычная гипотеза бесконтрольного накопления мутаций. Это, кстати, должно касаться не только морфологической редукции, но и фиксации модификаций: в этом случае мы имеем дело с процессом редукции морфогенетических механизмов (креодов и переключателей между ними), ответственных за выбор и осуществление альтернативных путей развития.
Для нас все же более существенна сама трудность перестройки однажды сформированного компромисса, существование лишь с трудом преодолимых барьеров на пути изменения сложившейся организации. По-видимому, эволюционный ландшафт не следует представлять так, как его обычно изображают: в виде адаптивных пиков, разделенных долинами пониженной приспособленности и соединенных хребтами, по которой эволюционирующая группа может постепенно двигаться по направлению от исчезнувшего пика к сохранившемуся. Более правдоподобной выглядит модель Ф.Р. Шрама (Schram, 1983) - система лунок, разделенных барьерами адаптивной неустойчивости.
Трудность преодоления устойчивости однажды достигнутого удачного компромисса предопределяет многие важные свойства эволюционного процесса: уже упоминавшиеся неравномерность и заторможенность эволюции, дискретность живых существ, малую эволюционную эффективность элиминации. Многие из вопросов, на которые синтетическая теория не может дать вразумительного ответа, такие как неравномерность эволюции, широкое распространение эволюционного стазиса, отсутствие видимого влияния популяционно-генетических факторов на скорость эволюционного процесса - все эти эффекты оказываются абсолютно естественными и ожидаемыми при рассмотрении процесса с позиции адаптивного компромисса. Однако, как и следовало ожидать, их место занимают другие проблемы, не менее трудные для разрешения.
Поскольку, несмотря на все барьеры, эволюция все же идет, хотя и ценой вымирания множества групп, не сумевших эти барьеры успешно преодолеть, попытаемся теперь понять, в каких условиях и каким образом один сбалансированный эпигенотип может быть преобразован в другой. Для этого проанализируем реальные условия, в которых такая перестройка идет, причем идет относительно быстро, в надежде обнаружить факторы, облегчающие искомые изменения.
Хотя репродуктивная изоляция не является необходимой для эволюции и даже не стимулирует ее (Гриценко и др., 1983, гл. 7), хорошо известно, что в условиях изолированных островов, водоемов и т. д. эволюция идет особенно быстро и часто приводит к образованию сильно измененных, даже гротескных форм. С.С. Шварц (1980), специально анализировавший этот парадокс, пришел к выводу, что причина состоит в пониженной напряженности конкурентных отношений бедных, незаполненных (в эволюционном масштабе времени) островных биоценозах. Конкретно эволюционная роль незаполненности ценозов состоит, по Шварцу, в том, что ослабленная конкуренция делает возможной быструю одностороннюю специализацию. Этот вывод находится в полном согласии с концепцией адаптивного компромисса: предковый вид, попадая с материка в обедненный островной биоценоз, оказывается в условиях, смягченных по многим параметрам и допускающих дополнительную оптимизацию функций, оставшихся под жестким контролем среды, за счет других, не контролируемых отбором столь же строго.
Изложенная модель эволюции на островах интересна тем, что она одновременно соответствует концепции инадаптивной эволюции по В.О. Ковалевскому (Расницын, 1986). Но еще более важно, что она демонстрирует достаточно реальный механизм, способный преодолеть, сломать устойчивость хорошо сбалансированного адаптивного компромисса, тормозящего эволюцию. Естественно возникает вопрос, не может ли этот механизм действовать не только при заселении островов, но и в каких-то других ситуациях.
Чтобы ответить на этот вопрос, рассмотрим некоторые другие случаи перехода группы в относительно мягкие условия - не будут ли и там наблюдаться ускорение эволюции и ее сходство с инадаптацией, т.е. увеличение размаха изменений в сочетании с их односторонностью, несбалансированностью. Одним из таких случаев может быть переход в новую экологическую нишу (точнее - создание новой ниши большой емкости, так как не занятых ниш, как известно, не существует). Поскольку каждый таксон - это и своя экологическая ниша (Шварц, 1980), можно допустить, что некоторое временное смягчение условий характерно для ранних (сразу за завоеванием новой ниши) этапов эволюции таксона. Тогда, если наше предположение верно, на ранних этапах эволюции таксона следует ждать, во-первых, ускорения эволюции (как частоты, так и масштаба изменения), во-вторых, интенсивного отмирания вновь образующихся инадаптивных групп по мере заполнения ниши.
Хорошо известный палеонтологам факт массового вымирания и диверсификации, часто происходящих почти одновременно, причем периоды диверсификации следуют за периодами вымирании (Грант, 1980, рис. 32, 5), подтверждает, что освобождение экологического пространства вымершими группами стимулирует эволюцию сохранившихся. Ускорение эволюции на ранних этапах истории таксона более конкретно демонстрирует факт приблизительного совпадения возраста рода и многих его видов (Шварц, 1980); это показывает, что значительная часть дивергенции происходит на самых ранних этапах эволюции возникшего таксона. Закон архаического многообразия (Мамкаев, 1968) фиксирует резко повышенную изменчивость организации на ранних этапах эволюции таксона, отчего виды и роды в это время могут различаться по признакам, позже характеризующим семейства и отряды. Не менее характерна для этапа архаического многообразия и малая эволюционная устойчивость ранних членов таксона, отражающаяся в обилии коротких базальных ветвей едва ли не на любой филогенетической схеме, построенной с использованием богатого палеонтологического материала.
Все это позволяет сделать вывод, то переход группы в более мягкие условия обедненного биоценоза и незаполненной экологической ниши действительно провоцирует эволюцию, причем эволюцию существенно инадаптивную. В таком случае становление эвадаптивных групп, т.е. гармоничных, приспособленных к заполненным биоценозам и напряженным конкурентным отношениям, должно быть связано с последующим ужесточением условий и сопровождаться интенсивным вымиранием большинства вновь возникших групп, именно тех, которые оказались неспособными достаточно быстро превратиться из инадаптивных в эвадаптивные.
Следующий вопрос, естественно возникающий из сделанного выше вывода - можно ли предполагать, что только смягчение условий способно провоцировать эволюцию. В этой связи проанализируем знаменитые опыты Г. X. Шапошникова (1961, 1965, 1966, 1978), в которых пересадка тлей Dysaphis anthrisci maicopica Shap. с пригодного кормового растения Anthriscus nemorosa MB. сначала на малопригодное (Chaerophyllum bulbosum L.), а затем на ранее совсем непригодное (Ch. maculatum Wild.) за считанное число поколений привела не только к значительным морфологическим изменениям, но и к утрате репродуктивной совместимости со своим видом и появлению неполной совместимости с D. chaerophyllina Shap., исконным потребителем Ch. maculatum.
Условия этого опыта трудно назвать мягкими. Тем не менее здесь обнаруживается важное сходство с эволюцией, спровоцированной смягчением условий - тот же односторонний, несбалансированный (инадаптирующий) характер эволюционно компетентного отбора. Действительно, в опыте способность питаться на новом растении-хозяине на какое-то время оказалась единственным жизненно важным комплексом адаптации, а все остальные отошли на задний план. Конечно, опыт не был завершен, но если бы он был продолжен и перенесен в природу, то мы, вероятно, смогли бы наблюдать и следующий этап эволюции: испытание зарождающейся группы всесторонним, эвадаптирующим отбором, для которого важны все аспекты адаптации - питание, размножение, индивидуальная устойчивость разных стадий онтогенеза в условиях различной плотности популяции, конкурентоспособность и т. д. Немного шансов, что конкретная популяция выдержит это испытание, но они есть.
Результаты опытов Шапошникова интересны и важны, но поставленный вопрос все еще не получил ответа. Можно ли считать эти условия жесткими? Смертность личинок колебалась на разных этапах эксперимента от 15-22 до 53-75%, что при указанном значении плодовитости (36,6 личинок от одной партеногенетической самки; данные только для конечного этапа опыта) означает выживание как минимум около 10 личинок в потомстве каждой самки и, следовательно, почти десятикратный прирост популяции в каждом поколении. Другими словами, смертность была весьма умеренной, очевидно, из-за того, что заботой экспериментатора враги тлей и другие неблагоприятные воздействия были по возможности устранены. Не удивительно, что линии в эксперименте оказались весьма устойчивыми и смогли быть поддержаны в течение 50 поколений - срок, который кажется не очень вероятным для отдельных линий в природных условиях. Что же до самого факта пересадки на малопригодное и совсем непригодное растение, то он как раз не является чем-то необычным. В природе тли должны часто оказываться в подобном положении: то крылатую самку занесло ветром в неподходящий биотоп, то дождь или ветер сбросили бескрылую тлю на другое растение. Просто в природе это трудно заметить, потому что там, в отличие от лаборатории, насекомое не на своем месте обычно гибнет.
Мы редко задумываемся, говоря о жестких, катастрофических и тому подобных условиях. Равновесная плотность популяции соответствует тем ее значениям, при которых смертность уравновешивает репродуктивный потенциал популяции, и в потомстве каждой самки удается оставить потомство в среднем только одной из всех ее дочерей ("закон Бекетова": А.Н. Бекетов, 1860; годом раньше об этом же писал, например, Ч. Дарвин, 1991: с. 69). Поэтому элиминация в популяциях в норме (за исключением кратких и более или менее уравновешивающих друг друга эпизодов колебаний численности) оказывается практически равной плодовитости, т.е. близкой к максимуму. Но если практически полная элиминация потомства есть норма существования популяции, если шансы на оставление потомства для каждой особи всегда минимальны, если вымирание целых популяций - обычное явление даже в экологическом масштабе времени, то что мы можем иметь в виду, говоря об ухудшении условий существования?
Похоже, что вопреки предыдущим рассуждениям ужесточения условий на популяционном уровне в природе вообще не бывает, и даже в приложении к видовому и надвидовому уровню о нем говорить трудно. Здесь ухудшение условий может означать лишь тривиальное (в эволюционном масштабе времени) вымирание видов, влияние которого на эволюцию будет ощущаться лишь как освобождение экологического пространства и, соответственно, смягчение условий. Но природа не терпит пустоты, а экологическое пространство особенно, и выжившие виды мгновенно заполнят пустоту, интенсифицировав процесс размножения сохранившихся популяций, и тем самым сведут к нулю кратковременное смягчение условий.
Таким образом, эволюционно значимое смягчение условий существования популяций, видов и других таксонов столь же сомнительно, как и их ужесточение. Смягчение условий само по себе означает лишь повышение выживаемости и, следовательно, рост популяции. В результате мальтусова геометрическая прогрессия размножения за считанное число поколений приведет к насыщению экологического пространства и восстановлению нормальных для особи и популяции условий, включая нормальную (равновесную с плодовитостью, т.е. почти стопроцентную) смертность. Изменение характеристик популяции в результате колебаний ее плотности ("волн жизни"), как известно, ограничено лишь изменениями в рамках нормальной внутривидовой изменчивости. Тот факт, что колебание плотности популяции представляет собой самое заурядное явление, и тем не менее случаев порожденного им необратимого эволюционного изменения до сих пор не описано, служит основанием для вывода о несущественности этого явления для эволюции.
Итак, ни смягчение, ни ужесточение условий существования не представляют в действительности реальных факторов эволюции. Что же в таком случае может преодолеть устойчивость сбалансированного эпигенотипа и успешно вывести организацию живого существа за пределы исторически обусловленной нормы?
Как мы установили, причина устойчивости сбалансированного эпигенотипа заключается в однажды достигнутом компромиссе между противоречивыми потребностями оптимизации разных адаптивных функций организма. Эти потребности противоречивы в том смысле, что из-за сложного переплетения морфогенетических и функциональных связей между всеми частями и признаками организма адаптивное усиление одной функции отзывается потерями эффективности других. Ослабление целостности онтогенеза, облегчающее независимые преобразования отдельных систем, конечно, не только возможно, но и имеет место в природе. Однако это направление (упрощение организации и общая дегенерация) достаточно специфично, требует особых условий и не является ни характерным, ни особенно широко распространенным. В конце концов, чтобы терять высоту организации, надо сначала ее приобрести. Ослабление целостности как механизм эволюции существует, но для объяснения эволюционного процесса его явно недостаточно.
Таким образом, нам необходимо реконструировать условия, в которых изменение организации происходит в условиях сохраняющейся (или растущей) целостности, либо, скорее, когда временное снижение целостности затем быстро восполняется. Для этого попытаемся уточнить метафору адаптивного компромисса. Главную проблему для эволюции представляет, как мы видели, противоречивый характер отношений между адаптивными функциями в процессе их оптимизации. Это противоречие создается, с одной стороны, тесными онтогенетическими и функциональными связями между системами обеспечения этих функций, с другой, жестким контролем за должным исполнением всех этих функций. Мы видели, что ослабление связей возможно, но недостаточно для объяснения эволюции. Общее ослабление селективного контроля за надлежащим исполнением адаптивных функций, как мы видели, тоже невозможно из-за того, что в эволюционном масштабе времени элиминация всегда приближена к максимально возможным значениям. Где же выход?
Подсказкой здесь служат относительно хорошо изученные случаи быстрой эволюции, в частности уже обсуждавшиеся ситуации, провоцирующие быструю эволюцию - островные условия и эксперименты Шапошникова. Общим для них является попадание популяции в новые, непривычные для них условия. То же предполагается и для взрывной эволюции при проникновении группы в новую экологическую нишу. Наиболее прозрачные для нас опыты с тлями показывают, в чем именно состоит необычность новых условий: в том, что экспериментатор предъявляет тлям очень жесткие требованию по одному или немногим параметрам (в данном случае - способность питаться на неподходящем растении), но при этом освобождает их от жесткого средового контроля по другим параметрам: защищает от непогоды, хищников, паразитов и болезней, и т.д.
Мне неизвестны исследования, анализирующие подобный расклад интенсивности селективного контроля за разными адаптивными функциями у вселенцев на острова или в новые экологические ниши. Однако общие соображения о низкой, по сравнению с материковыми, напряженности конкурентных отношений в островных условиях весьма правдоподобны и подтверждаются относительно высоким иммиграционным потенциалом материковых видов на островах и крайне низким в обратной ситуации. В таких условиях ценотический контроль за исполнением некоторых функций (таких, как, например, функции защиты от хищников и специализированных паразитов, которые в островных сообществах нередко в дефиците) должен оказаться пониженным за счет усиления контроля по другим направлениям. В конце концов, на то условия и новые, чтобы расклад селективных требований там был иным, чем прежде, и в частности, чтобы некоторые функции контролировались существенно жестче, чем прежних условиях. Если же популяция тем не менее выжила, значит, суммарная элиминация там устойчиво не превышала репродуктивный потенциал и, следовательно, селективный контроль других функций был снижен.
Таким образом, эволюцию провоцирует, по-видимому, только одностороннее смягчение средового контроля организации, при котором лишь некоторые из адаптивных функций остаются под жестким контролем. Очевидным следствием такого одностороннего (несбалансированного) контроля является разбалансировка, дестабилизация сложившейся организации, и она действительно проявляется в подобных случаях (Жерихин в Деятельность..., 1967; Беляев, 1974; Шишкин, 1984, 1987; McCune, 1990, и др.). Поскольку при дестабилизации адаптивность системы неизбежно снижается, перестройка едва ли может происходить в нормальных условиях. Из этого следует, между прочим, что нарождающаяся группа вряд ли может внедриться в уже оккупированную нишу и вытеснить ее владельца. Более правдоподобно, что она либо займет нишу, освободившуюся при вымирании ее прежнего обитателя, либо сумеет создать (открыть) новую нишу. Может быть, именно поэтому нам известно довольно много правдоподобных примеров недавнего возникновения новых таксонов в островных условиях, а также при интродукции - иногда даже в историческое время. Особенно поражают гельминтологические примеры (цитированные в дискуссии по докладу Rausch, 1982: 184) о двух последовательных актах видообразования за 300 лет (у паразита домовой мыши, интродуцированной в С. Америку) и о дивергенции двух человеческих лентецов не более 16 тыс. лет назад. А вокруг нас, вокруг нас, где и биоразнообразие, и степень его изученности гораздо выше, и стало быть вероятность обнаружения случаев естественного видообразования гораздо больше, таких примеров очень немного.
Сказанное позволяет высказать в качестве рабочей гипотезы предположение, что эволюция на видовом и в особенности на более высоких таксономических уровнях возможна лишь в результате такого изменения условий, при котором действующий на популяцию отбор становится существенно односторонним. Только такой отбор представляется способным преодолеть устойчивость прежнего хорошо сбалансированного адаптивного компромисса. Однако вновь возникшая группа, прошедшая дестабилизацию и потерявшая былую сбалансированность, должна пройти жесткий всесторонний контроль (отбор по всем адаптивным функциям) и под его действием восстановить свою устойчивость. Другими словами, эволюционное изменение должно пройти через два этапа - дестабилизацию и затем стабилизацию сбалансированного эпигенотипа.
Как уже упоминалось, результаты этих событий можно отождествить соответственно с инадаптивным и эвадаптивным изменением организации. Другими словами, в процессе эволюции организация живых существ проходит три последовательных этапа: инадаптацию, эвадаптацию и стазис. Если, конечно, не случится вымирания, что возможно на любом из этапов, хотя наиболее рискованным в этом плане, естественно, является процесс эвадаптации, а наиболее спокойным - стазис. Первые два этапа как более уязвимые по необходимости должны быть кратковременными, а стазис может продолжаться неопределенно долго. Выход из стазиса едва ли может быть спонтанным: причиной должно стать изменение условий. Если такое изменение вызывает рост общей элиминации за пределы репродуктивного потенциала, неизбежно вымирание популяции (и вида, если вымерли все его популяции, или старшего таксона, если то же случилось со всеми его видами). Если же условия существования меняются таким образом, что контроль организации становится односторонним, не исключено, что популяция сможет войти в процесс инадаптивного преобразования ее сбалансированного эпигенотипа. А вот сумеет ли она успешно пройти его, и тем более - пройти крайне рискованный процесс эвадаптации, это уже дело случая, причем случая весьма редкого.
Здесь нужно отметить еще одну важную черту нашей модели - ее авторегуляторность. Сколь высоко ни поднялась бы устойчивость (забуференность) эпигенотипа даже заметной части обитателей нашей планеты, даже большие катаклизмы вряд ли будут серьезно угрожать разнообразию жизни на Земле, если, конечно, иметь в виду эволюционно значимые отрезки времени. Дело в том, что даже в том случае, когда заметная часть биоразнообразия, "закосневшая" в совершенстве и, стало быть, жесткости своих эпигенотипов, окажется существенно "прореженной" очередным субглобальным оледенением, астероидной либо ядерной зимой или подобным апокалипсисом, выжившие виды попадут в ситуацию неполных сообществ с существенно ослабленными конкурентными отношениями. Это будет провоцировать быструю инадаптивную эволюцию, и тем более быструю, чем более обширной была "прополка". По мере заполнения сообществ многие из вытащивших выигрышный билет его потеряют (ср. модель Flush and Crash: Carson, 1968), но не все: чем больше доля неудачников, тем больше шансов у тех, кто сумел продержаться чуть дольше их, и тем дольше будет процесс восстановления былого разнообразия. Свято место пусто не останется. Правда, наш оптимизм не распространяется так далеко, чтобы обещать новые сообщества похожими на прежние. Как известно, вымирание даже в окрестностях границы перми и триаса было хотя и масштабным, но не подавляющим: обычно говорят о потере с середины верхней перми порядка половины семейств и трех четвертей родов в морях (см., например, Алексеев и др. 2001, рис. 6б,в) и существенно меньше на суше (там же, рис. 25б, 32а, 42а). Тем не менее мир после этого, оправившись приблизительно к середине триаса, оказался очень существенно иным, причем не только по составу, но и по тенденциям. Так, суммарное разнообразие, достигнутое в морях во второй половине ордовика, далее до середины верхней перми (двести миллионов лет, т.е. лишь немного меньше, чем мезозой и кайнозой вместе!) не росло и лишь колебалось вокруг позднеордовикского уровня. После же пермотриасового кризиса рост возобновился и продолжается до сих пор, уже многократно перекрыв позднепалеозойский уровень биоразнообразия (Алексеев и др. 2001, рис. 6). Очевидно, сама структура морских сообществ радикально изменилась, дав место гораздо большему росту биоразнообразия.
Другая важная черта нашей модели эволюции - относительно малая предсказуемость результатов ее действия (весьма неполное соответствие между условиями протекания эволюции и ее результатами), что обусловлено большой сложностью эволюционирующей системы и, соответственно, невозможностью простых, предсказуемых решений. Это, конечно, огорчительное свойство модели, но в условиях, когда предсказуемость эволюционного процесса, т.е. корреляция между признаками организации живых существ и условиями их существования, в действительности очень невелика, это и важное ее достоинство. По крайней мере по сравнению с синтетической теорией эволюции, предсказывающей прогнозируемость эволюции.
Подход к адаптации как к компромиссу позволяет привести в соответствие с селекционистской парадигмой ("эволюция есть процесс взаимодействия отбора с наличной организацией, т.е. с продуктом отбора предшествующих поколений") еще одну важную закономерность эволюции, не выводимую в общем виде из синтетической теории. Я имею в виду неравномерность эволюционного процесса, столь неопровержимо вытекающую из палеонтологических данных. Трехчленный цикл "инадаптация - эвадаптация - стазис" с очень быстро протекающими двумя первыми этапами и длительным периодом эволюционной стабильности легко находит параллели с моделями, предложенными палеонтологами (Симпсон, 1948; Eldredge, Gould, 1972; Stanley, 1975; Gould, Eldredge, 1977) и генетиками (Carson, 1975) и нацеленными на введение неравномерности в "синтетическую" модель эволюции. Это сходство не следует переоценивать, так как сходна лишь феноменология, а предполагаемые механизмы неравномерности в разных моделях различны, в том числе и в отношении их реалистичности. В рамках синтетической теории эволюции трудно предложить для эволюционного стазиса иные механизмы, кроме нивелирующего эффекта обмена генами и стабильности условий. Это не значит, конечно, что сторонники синтетической теории никогда не обращались к идее устойчивости сбалансированного эпигенотипа. Именно об этом писали Э. Майр (1974; см. выше) и другие авторы (например, Van Valen, 1982; Laurin, Bruno, 1988). Проблема только в том, что эта идея из совершенно другой, не синтетико-эволюционной парадигмы.
Таким образом, можно заключить, что многие наблюдаемые особенности эволюционного процесса находятся в неплохом соответствии с предсказаниями эпигенетической теории эволюции и в частности метафоры адаптивного компромисса. Однако остаются и проблемы. Загадка распределения эволюционных скоростей, столь очевидно противоположного предсказаниям синтетической теории, пока не поддается безупречному объяснению и с помощью эпигенетической теории, хотя и не находится в столь же неустранимом противоречии с ней.
Действительно, имеющиеся данные прямо противоречат концепции адаптивного компромисса, по крайней мере на первый взгляд. Если основным тормозящим эволюцию фактором мы считаем компромиссную природу адаптации, и в частности трудность изменения хорошо сбалансированного эпигенотипа, то с усложнением эволюционирующей системы и, следовательно, ужесточением компромисса следовало бы ожидать усиления этого торможения. На самом же деле, как мы видели, эволюция более сложных форм жизни протекает в целом быстрее.
Не все, однако, так просто. Прежде всего, повышенная трудность изменения .высших организмов, предсказываемая концепцией адаптивного компромисса, означает меньшую частоту возникновения у них адаптивных изменений, но ничего не говорит о других параметрах, определяющих скорость эволюции. Важным фактором, который может избирательно увеличивать скорость эволюции высших групп организмов является, по-видимому, направленность изменений, своего рода ортогенетический фактор, повышающий кумулятивный эффект ряда последовательных изменений. Действительно, при ненаправленном, хаотическом характере внешних изменений, благоприятствующем броуновской эволюции, случайный возврат условий в состояние, близкое к существовавшему ранее, будет воспринят как таковой лишь организмами, которые за время, разделяющее эти сходные условия, изменились поверхностно, неглубоко. При более глубоком, целостном преобразовании организации возврат к объективно прежней ситуации измененная система субъективно воспримет как совершенно новую обстановку, требующую соответствующих, т.е. новых, адаптаций. В результате эволюция высших форм жизни будет включать меньший броуновский компонент по сравнению с низшими, что должно увеличивать эффект суммы последовательных изменений за длительное время (Alberch, 1980; Rachootin, Thomson, 1981; Oster, Alberch, 1982).
Существенно также, что напряженность компромисса, затрудняющая его перестройку, определяется не только внутренними факторами (степенью взаимовлияния систем обеспечения различных функций), о чем шла речь выше. Не менее важны жесткость и особенно всесторонность внешнего (средового и преимущественно ценотического) контроля над адаптивностью системы, определяющие возможность изменения одних ее подсистем ценой временного снижения адаптивности других. Правда, наше неумение сравнивать уровень внешнего контроля у разных групп исключает на современном этапе возможность прямо оценить его влияние на распределение темпов эволюции. Однако нам остаются обходные пути, например, сравнение общей устойчивости разных организмов и их групп к изменениям условий, так как эволюционирующая система воспринимает эти изменения именно как изменения контроля над адаптивностью отдельных функций.
Мы можем, например, попытаться сравнить характер индивидуальной и популяционной устойчивости в группах разного эволюционного уровня. Количественным показателем индивидуальной устойчивости, в соответствии с "законом Бекетова" является характерный для группы темп воспроизводства: плодовитость и длительность генерации. Совершенно очевидно, что темп воспроизводства в целом падает, а индивидуальная устойчивость растет с повышением уровня организации. Соответственно растет и степень автономизациии организма от среды, что, казалось бы, должно вести к повышению и эволюционной устойчивости, т.е. к замедлению эволюции высших форм. Дело, однако, в том, что автономизация весьма энергоемка, поскольку требует более тонкой и быстрой оценки состояния среды, без которой невозможны упреждающие реакции на неблагоприятные изменения. Это ведет к повышению зависимости организма от источников пищи. Как пишет С.А. Северцов, "горностай, которому для насыщения нужно в день добыть количество мяса, равное половине его веса, и гадюка, которая полгода проводит в оцепенении, а в летний период довольствуется двумя-тремя полевками в неделю, представляют хороший пример различия в энергии жизнедеятельности. Крокодилы в зоологическом саду довольствовались 36 г мяса в день. Близким по весу пантерам и леопардам дают 3,5-4 кг мяса в день" (1941, с. 288).
Механизмы автономизации рассчитаны на нормальные, привычные для вида изменения условий, они оппортунистичны, как все адаптации, и их совершенство, достигаемое ценой повышенной потребности в энергии, отнюдь не гарантирует более высокой эволюционной устойчивости к новым для вида условиям.
Обратимся теперь к популяционному механизму эволюционной устойчивости. Этот механизм - избыточная (резервная) плотность популяции, вызывающая повышение смертности и (или) снижение плодовитости по зависимым от плотности факторам, влияющим на численность популяции (Van Valen, 1976). Популяционная устойчивость универсальна, поскольку снижение избыточной плотности одинаково компенсирует снижение средней выживаемости независимо от того, чем это снижение вызвано. Однако объем резервной плотности ограничен, так как повышение плотности популяции невозможно за пределы емкости пространства ресурсов, а "срабатывание" избыточной плотности возможно лишь до определенного уровня, ниже которого выживание разреженной популяции становится проблематичным.
Значение резерва избыточной плотности популяций для эволюции демонстрирует описанный С.В. Мейеном (1987а) эффект фитоспрединга, согласно которому новые таксоны высокого ранга возникают в экваториальной зоне, с климатическими колебаниями проникают в более высокие широты и частью, адаптируясь, задерживаются там (сходные мысли ранее высказывал Дарлингтон, 1966; обобщение этой модели предложил Eskov, 2002, но сейчас это нам не столь важно). Для объяснения фитоспрединга я предположил (Расницын, 1989г), что дело здесь именно в резервной плотности популяций. Действительно, условия жизни и суммарное давление на популяцию в разных местах на Земле (кроме островных условий) не может существенно и устойчиво различаться: миграции и эволюция этого не допустят и все "свободные" места будут быстро (в эволюционном масштабе времени) заполненены. Но по крайней мере абиотические условия в несезонном климате в целим мягче, чем в сезонном, и эта разница должна быть компенсирована большей напряженностью биотических и прежде всего конкурентных отношений, включая и внутривидовых, что соответствует повышению резервной плотности популяции. При прочих равных это делает возможными более глубокие эволюционные изменения в тропиках по сравнению с умененным и холодным климатом.
Для нашей проблемы, однако, важно другое - имеется ли корреляция между объемом резервной плотности популяции и уровнем организации. Прямое численное сравнение избыточной плотности, скажем, у бактерий и млекопитающих кажется рискованным, и мы снова изберем обходной путь анализа факторов, влияющих на эту плотность и одновременно зависящих от эволюционного уровня. Объем избыточной плотности в целом определяется положением порогов устойчивой плотности - нижнего, за которым популяция вымирает из-за слабости необходимого для выживания взаимодействия особей, и верхнего, превышение которого невозможно из-за ограниченности ресурсов.
Верхний порог устойчивой плотности в конечном счете определяется величиной доступного популяции потока энергии, который она может использовать в обеспечение ее существования. При постоянстве этого потока верхний порог устойчивой плотности зависит от энергетических потребностей особи (поскольку обсуждаемые ниже факторы, влияющие на нижний порог устойчивой плотности, зависят от плотности особей, а не их суммарной биомассы, то и здесь приходится оперировать тем же показателем). Энергетические же потребности особи быстро растут с повышением организации, во-первых, из-за в среднем более крупных размеров высших организмов, во-вторых, за счет дополнительных затрат на автономизацию, о которых уже говорилось. Таким образом, рост энергозатрат особи с повышением уровня ее организации, и соответствующее снижение верхнего порога устойчивой плотности популяции должны расти весьма быстро.
Нижний порог устойчивой плотности возникает (или повышается), например, с появлением полового процесса, точнее, связан с необходимостью встречи двух особей, причем не любых двух, для успешного размножения. Сходный эффект дают и адаптации кондиционирования среды (ее изменения в благоприятном направлении деятельностью группы особей). Первый фактор явственно, хотя и не жестко связан с высотой организации (партеногенез и бесполое размножение наблюдаются и в довольно продвинутых группах организмов), для второго такая связь не очевидна, но допустима, поскольку высокая организация благоприятствует развитию более эффективных форм взаимодействия. В итоге можно предположить, что повышение организации сопряжено с определенным повышением нижнего порога устойчивой плотности популяции и, следовательно, с некоторым снижением устойчивости к неблагоприятным воздействиям. С учетом же сильной зависимости верхнего порога от высоты организации можно с уверенностью заключить, что объем резервной плотности популяции находится в обратной связи с высотой организации, и это не может не снижать эволюционную устойчивость и, соответственно, повышать скорость эволюции высших групп организмов.
Далее, устойчивость популяции и вида в изменяющихся условиях также в немалой степени зависит от того, насколько организмы способны найти в новой среде мало изменившиеся островки и убежища и удержаться в них. В большой степени это определяется расселительными способностями организмов. Действительно, при анализе данных по темпам эволюции (Расницын, 1987, табл. 1, 2) бросается в глаза относительно низкая скорость эволюции птиц (возраст полусовременной фауны на уровне отряда, т.е фауны, в которой половина отрядов вымершие, половина современные, у птиц 100 млн. лет против 70 у рептилий и 40 и млекопитающих; время полувымирания видов рукокрылых 1,6 млн. лет против 0,18-0,49 у других млекопитающих). Однако в целом прямая связь расселительных способностей с высотой организации, если и существует, вряд ли велика.
Более важны в этом плане косвенные факторы и в частности роль абсолютных размеров организма: там, где мелкие формы могут найти себе мало изменившиеся микробиотопы, крупные часто оказываются вынужденными измениться или вымереть. Так, возраст полусовременной фауны крупных и мелких млекопитающих, как уже упоминалось, различается в 2,5 раза (200 тыс. лет для слонов и копытных против 500 тыс. лет для грызунов, насекомоядных и летучих мышей, Расницын, 1987, табл. 1), причем эти даты приходятся на крайне изменчивый по условиям плейстоцен. У насекомых полусовременная фауна по тем же данным датируется приблизительно 5 миллионами лет. Большая часть крупных млекопитающих из своеобразной фауны вымершего тундростепного ландшафта, занимавшего в четвертичное время обширные пространства севера Евразии, как известно, вымерла в начале голоцена, а мелкие в основном выжили (Шер, 1997а,б; Жегалло и др., 2001). Насекомые же оказались устойчивыми в течение всего ледникового периода и послеледниковья: по данным Назарова (1984), из 2000 видов насекомых, известных для четвертичного периода, вымершими считаются не более 30. Более того, значительная часть характерных тундростепных видов насекомых позднего плейстоцена сохранилась не только в степях Южной Сибири и Монголии, но и в местах своего прежнего обитания - на небольших реликтовых "степных" участках в Якутии и даже на Чукотке (Берман, 2001).
Размеры определенно, хотя и не строго, связаны с высотой организации: малые размеры ограничивают возможность усложнения, а крупные требуют какой-то минимальной сложности хотя бы для жизнеобеспечения и координации действий удаленных частей организма. Поэтому снижение эволюционной устойчивости высших форм жизни из-за меньшей, по сравнению с низшими, доступности для них островков стабильности в измененных условиях, должно весьма заметно снижать их эволюционную устойчивость и увеличивать скорость как вымирания, так и эволюционных преобразований.
Перечисленные выше факторы вряд ли исчерпывают все разнообразие связей между высотой организации и устойчивостью, определяющее темпы эволюции, однако другие механизмы пока не столь очевидны. Так что в итоге нам удалось обнаружить один механизм, который, если концепция адаптивного компромисса верна, должен снижать темпы эволюции высших форм жизни. Это компромиссная природа адаптаций как таковая, т.е. трудность изменения более сложной сбалансированной системы. В противоположном направлении действуют несколько факторов, коррелированных с усложнением организации. В их числе:
1) сильнее выраженная направленность их эволюции,
2) снижение эволюционной устойчивости более сложных организмов, обусловленное сближением порогов устойчивой плотности, а именно:
а) понижение верхнего порога устойчивой плотности из-за дополнительных энергозатрат, во-первых, на более крупные размеры их особей, во-вторых, на обеспечение опережающего роста их механизмов автономизации от внешней среды,
б) повышение нижнего порога устойчивой плотности популяции с появлением полового размножения (из-за необходимости обеспечить встречу полов,
3) меньшая доступность для более сложных организмов "островков стабильности", сохраняющихся при общем изменении обстановки, из-за связи между высотой организации и абсолютными размерами.
Конечно, хотелось бы получить данные о фактической эффективности этих факторов из независимых источников, чтобы оценить их относительное значение и интегральный эффект. Только тогда мы получим возможность судить с определенностью о том, насколько правдоподобно наше объяснение парадокса скоростей эволюции: действительно ли совокупность именно этих факторов обеспечивает ускорение эволюции эволюционно продвинутых групп организмов. Тем не менее уже сейчас можно сделать вывод, что гипотеза адаптивного компромисса как часть эпигенетической теории эволюции неплохо согласуется с накопленными данными по феноменологии эволюционного процесса на уровне организма и таксона. В том числе и с теми наблюдениями, интерпретация которых вызывает серьезные трудности для синтетической теории.
Завершая раздел по онтологии эволюционного процесса, рискну сделать вывод, что эпигенетическая теория эволюции находится в хорошем согласии не только с результатами наблюдений и экспериментов в области осуществления и преобразования онтогенеза, но и с палеонтологическими данными по эволюции организмов. Уверенной интерпретации оказался доступен довольно большой массив фактических данных в обеих областях, и это вселяет надежду, что и оставшиеся проблемы и вопросы со временем будут успешно разрешены.
МЕТОДОЛОГИЯ ФИЛОГЕНЕТИКИ, ТАКСОНОМИИ И НОМЕНКЛАТУРЫ
ЭЛЕМЕНТЫ ОБЩЕЙ МЕТОДОЛОГИИ
В своей обычной работе ученый не фиксирует внимание на ее методологических основах. Однако когда привычные стандарты и методы начинают давать сбои или просто перестают нас удовлетворять, возникает соблазн устроить нечто вроде научной революции (Kuhn, 1970): отбросить привычные методы и стандарты вместе с методологией, на которой они базируются, и заменить чем-то другим. Революции, даже научные, редко проходят бескровно, в том смысле, что они отбрасывают подходы, еще вполне работоспособные, хотя и в несколько ином контексте, и результаты значимые, но требующие переосмысления. Поэтому предпочтительнее заново проанализировать методологические основы научной деятельности в надежде выявить и исправить слабые места и тем сохранить как сам подход, так и основную часть полученных с его помощью результатов.
Нынешняя революция в теории систематики (таксономии) и тесно связанной с ней филогенетике тянется уже полвека и порождена, на мой взгляд, стремлением к формализации, автоматизации и стандартизации научного процесса, к усилению его объективности и воспроизводимости результата. И соответственно к снижению роли индивидуальности исследователя, особенно его интуиции.
Люди старшего поколения помнят дискуссию о физиках и лириках, чью-то знаменитую строчку "Что-то физики в почете, что-то лирики в загоне" и новосибирского кибернетика Игоря Полетаева, призывавшего изгнать методы искусства из научного исследования. У нас эта революция началась бурными дискуссиями интеллектуалов, но быстро затормозилась из-за слабости технической базы и ограниченных связей нашей науки с западной.
А на Западе интенсивная компьютеризация влияла не только на быт, промышленность и исследовательскую технику, но и на психологию и идеологию ученого, и требовала формализованных, математизированных методов, свободных от интуиции или хотя бы кажущихся свободными. Первой фавориткой систематиков стала фенетика, потом их умами целиком завладела кладистика, благо давние труды Вилли Геннига были наконец открыты американцами и переведены на английский. Кладистика и поныне в основном соответствует западному, прежде всего американскому мироощущению, хотя и там уже возникают вопросы и скепсис. У нас же кладизм лишь начинает перерастать из моды в образ мыслей, и сравнительный анализ кладистической и не-кладистических методологий встречает меньшее сопротивление.
Шаги познания
Каждый шаг познания включает, на мой взгляд, шесть последовательных этапов (Табл. 1). Опираясь на предшествующий опыт и знание, мы ставим задачу и планируем сбор материала. Затем проводим наблюдения и/или ставим эксперимент и анализируем из результаты, пытаясь отыскать знакомые элементы в незнакомой (еще не понятой) видимой картине. Опираясь на общий опыт и результаты предыдущего знакомства с подобными объектами, мы выделяем среди знакомых элементов более важные для целей нашей работы, т.е. те, что могут помочь нам выявить важные черты пока непонятной картины. В свою очередь, важными мы считаем черты, оказывающие, как мы предполагаем, сильное влияние или иначе тесно скоррелированные со структурой нашей картины и тем позволяющие ее понять и предвидеть ее поведение в разных обстоятельствах. Например, когда систематик обнаруживает совершенно новую группу насекомых и начинает их классифицировать (создавать внутреннюю систему группы), то, зная о существовании и возможных различиях полов и стадий онтогенеза, он исключит признаки, различающие самцов и самок, взрослых и личинок и т. д., из числа таксономически важных признаков, как бы ни глубоки были эти различия сами по себе. Сходным образом мы обычно оказываем предпочтение структурным признакам перед признаками окраски.
Таблица 1. Шаги познания
|
1 |
постановка задачи и планирование сбора данных |
|
2 |
наблюдение и эксперимент |
|
3 |
поиск аналогий: знакомых элементов в новой для нас картине, и выделение более важных из них - тех, что могут отражать глубинную структуру системы |
|
4 |
создание гипотез о закономерностях и механизмах, лежащих в основе видимой картины |
|
5 |
попытка фальсификации этих гипотез: выведение следствий из них и их сравнение с результатами наблюдений и экспериментов, как прежних, так и новых, проведенных специально для проверки гипотез |
|
6 |
оценка результата с помощью презумпций: если найдены фальсифицирующие доказательства, нужно решить, достаточно ли они надежны и убедительны, чтобы отказаться от гипотезы |
Здесь начинается третий этап. Мы строим гипотезу об особом сходстве между исследуемой картиной и некоторым классом картин (patterns), изученных ранее и отличных от изучаемой. Гипотеза должна быть прогностичной, т.е. указывать на некий механизм, ответственный за сходство, или предсказывать сходство по многим другим признакам. Это позволит сделать дальнейшее предположение о поведении нашей картины (системы, совокупности предметов и явлений) в различных условиях и обстоятельствах. Возвращаясь к предыдущему примеру, систематик выдвигает гипотезу, что, например, сходство по структурным признакам указывает на более высокий (скажем, родовой) уровень общего сходства, тогда как окраска относится к более низкому (например, видовому) уровню. Другими словами, систематик предполагает, что эта гипотеза позволит классифицировать его группу насекомых более естественным образом, чем альтернативный подход (например, придание большего веса признакам окраски).
Три предыдущие этапа познания сильно зависят от личного опыта и интуиции исследователя. Их формальная когнитивная структура еще не ясна, и я не буду обсуждать их в деталях.
Четвертый этап - тестирование, проверка предложенных гипотез. Для этого создаются возможно более длинные и разветвленные цепи следствий, дающие больше возможностей для сравнения сделанных таким образом предсказаний с результатами наблюдений и экспериментов (подробнее см. ниже).
Пятый и последний этап состоит в принятии решения, приемлема ли каждая из предложенных гипотез по результатам испытания, следует ли ее сохранить для дальнейшего тестирования, или она должна быть отброшена. Опять-таки, примеры и обсуждение следуют далее.
Со времен Карла Поппера (1983) известно, что все результаты научного исследования - не более чем гипотезы, которые невозможно окончательно подтвердить (верифицировать). Поппер, правда, считал, что гипотезу можно хотя бы окончательно опровергнуть (фальсифицировать), но он был неправ, так как окончательная фальсификация одновременно есть окончательная верификация альтернативной гипотезы, что фальсифицирующий результат не есть ни случайность, ни результат действия неучтенных факторов и обстоятельств. Действительно, если вы утверждаете, что все лебеди белые, а вам предъявляют черного, то прежде чем отказаться от вашей гипотезы, следует попытаться отвергнуть по крайней мере две другие возможности: во-первых, что это не лебедь (а, например, гусь, или искусная подделка, или что-то еще в этом роде), во-вторых, что лебедь не по-настоящему черный (например, искусственно окрашен). По моему убеждению, эти альтернативные возможности всегда оцениваются в терминах вероятности (правдоподобия), а не простого ответа "да" или "нет", как считал Поппер. Именно поэтому оказывается необходимым пятый этап познания, когда мы сравниваем конкурирующие гипотезы и принимаем решение, оценивая альтернативные фальсифицирующие доказательства.
Так что все наши знания - это гора опирающихся друг на друга гипотез, причем гора вершиной вниз, поскольку базовых гипотез мало, а опирающихся на них частных - бездна. Но горе еще и в том, что наши инструменты анализа не соответствуют тому материалу, которым призваны оперировать: они ориентированы на поиск истины, а не на манипуляции с гипотезами. Доказательства отталкиваются от положений, принимаемых за истинные, но даже в математике эти базовые постулаты могут быть оспорены, а у нас в биологии их и вовсе не сыскать. Законы как эмпирические обобщения высокой степени надежности вовсе не так надежны - даже такие фундаментальные, как законы сохранения вещества и энергии. А наши законы, вроде биогенетического, необратимости или неспециализированного предка, весьма мало надежны. Объяснение как сведение наблюдаемых феноменов к процессам других уровней, которые далее не анализируются, т.е. принимаются за истинные, тоже не слишком надежный инструмент для проникновения в природу вещей. Во-первых, истинность результатов, полученных на других уровнях знания, не обязательно выше наших и во всяком случае не абсолютна. По-хорошему нужно продолжать анализ дальше, на все более высоких (или более низких) уровнях - до бесконечности.
Во-вторых (а может быть, во-первых), одни и те же эволюционные феномены объясняются, и нередко с приблизительно равным успехом, синтетистами, ламаркистами и теологами. Не случайно Поппер развил фальсификационистскую концепцию, отталкиваясь от своего фрейдистского опыта, когда убедился, что нет и, главное, невозможно себе представить ситуацию, которая не может быть успешно объяснена в рамках психоанализа. Именно на этом основании он отказал фрейдизму, а равно марксизму и дарвинизму в принадлежности к науке. Справедлива ли его оценка, вопрос отдельный. Мне кажется, Поппер не прав, поскольку в соответствии со своей концепцией фальсификационизма он имел в виду experimentum crucis: решающий эксперимент, который один может дать окончательный ответ на поставленный вопрос, фальсифицировав всю соответствующую систему взглядов. Как следует из предыдущего, я не считаю это возможным: как и отдельную гипотезу, так и их систему может опровергнуть, и то не окончательно, а с той или иной степенью правдоподобия, только большая совокупность наблюдений. Как это может выглядеть на практике, вопрос специальный, и решать его здесь я не берусь.
Как бы то ни было, получается, что мы обладаем только очень ненадежным знанием и неадекватными инструментами для работы с ним. Однако в этом пессимистическом выводе что-то явно не так: если бы он был справедлив, наука не достигла бы и сотой доли того, что достигла. Более того, в обыденной жизни истина нам еще менее доступна, чем в науке, а задачки там попадаются куда более ответственные, когда цена ошибки - жизнь, и порой жизнь многих. Тем не менее и эти задачи мы в целом решаем достаточно успешно, чтобы выживать при нашем ничтожном репродуктивном потенциале. Очевидно, адекватные инструменты у нас есть, и мы их активно используем, только не даем себе в этом отчет, как мсье Журден не догадывался, что говорит прозой. Вытащить этот инструмент из подсознания сумели, и очень давно, юристы, но и они не заподозрили его универсального значения. Этот инструмент, по моему убеждению, презумпция (Расницын, 1988в). Ее парадигмальный вариант (образец, с которым можно сверять все другие презумпции, т.е., говоря по-нашему, типовой экземпляр) - это, конечно, презумпция невиновности: "Как бы тяжелы ни были подозрения, подозреваемый должен считаться невиновным, пока и поскольку его вина не доказана должным образом". Что такое "должным образом", вопрос отдельный и очень важный, к нему еще придется вернуться. В остальном же все презумпции построены однотипно: для определенного класса задач презумпция предлагает типовое решение, которое следует принять независимо от того, есть ли или нет свидетельства в его пользу, но лишь тогда, когда отсутствуют надежные свидетельства противоположного свойства.
Вот несколько примеров презумпций из нашей области (подробнее о презумпциях из нашей области см. ниже). Последовательность слоев земной коры следует интерпретировать как временную последовательность напластований, пока и поскольку нет надежных свидетельств обратного. Гомотаксальные (сходного таксономического состава) комплексы ископаемых следует рассматривать как геологически синхронные, если тому нет серьезных контраргументов. Любой таксон нужно считать монофилетическим, если нет убедительных свидетельств его полифилии. Сходство организмов должно рассматриваться как унаследованное до тех пор, пока не будут представлены надежные аргументы его независимого происхождения. Онтогенетическую последовательность изменений организма следует интерпретировать как эволюционную последовательность, если только нет достаточных оснований интерпретировать ее иным образом. И вообще, не следует умножать сущности без нужды (бритва Оккама), т.е. принимайте самые простые решения, пока и поскольку нет серьезных оснований принимать более сложные.
На что я хотел бы обратить внимание. Презумпция есть не более чем концентрированный опыт удачных решений конкретных задач. Так в норме для здоровья общества опаснее осудить невиновного, чем оставить преступника на свободе. Нормальная последовательность слоев более обычна, чем обратная. В геологическом масштабе времени миграции обычно мгновенны, и существенная асинхронность сходных по составу комплексов ископаемых - вещь не частая. Доказуемая полифилия таксона - ситуация не слишком обычная, особенно у высших организмов. Совпадение, в первом приближении, онтогенетической и палеонтологической последовательностей действительно бросается в глаза во многих случаях. И т.д. Но всегда есть исключения, и порой их много. Так много, что применимость презумпции нередко бывает ограниченной. По утверждению Г.А. Заварзина (1987, см. также Woese, 1987), у микроорганизмов сходство (кроме как по первичной структуре ДНК) чаще приобретается независимо, чем наследуется, и соответствующая презумпция к ним неприложима. В четвертичной палеонтологии с ее дробной хронометрией миграции нередко фиксируются, вызывая асинхронность гомотаксальных комплексов. Советское общество было организовано таким образом, что его здоровье поддерживала скорее презумпция виновности (т.е. реально правосудие там реально вершил не суд, а следствие, чьим законным инструментом является презумпция виновности). И так далее.
Уже по одному этому не следует анализировать презумпции в терминах истинности или пригодности в целом: скорее следует говорить о том, что данная презумпция неплохо работает в данном контексте, а альтернативная ей успешно применяется в некоторых других случаях. Следует еще иметь в виду, что для решения одной и той же задачи могут одновременно применяться разные презумпции, и рекомендуемые ими решения могут оказаться несовместимыми. Так, относительно эволюционной последовательности изменений некоторой структуры (в филогенетической терминологии это задача поляризации признака) важные указания могут давать палеонтологическая и онтогенетическая последовательности, характер распространения разных состояний признака в исследуемой совокупности и за ее пределами, сложность и степень функционального совершенства разных вариантов данной структуры, наличие рудиментов и т.д. Каждому из этих подходов соответствуют свои презумпции, и результаты их приложения к конкретной задаче нередко противоречивы. Например, многие признаки черепа взрослого человека соответствуют эмбриональным признакам человекообразных обезьян, а в нашем онтогенезе соответствующих признаков взрослых человекообразных нет. Т.е. биогенетическая презумпция указывает на происхождение обезьян от человека. Напротив, в палеонтологической летописи черепа человека появляются значительно позже обезьяньих, свидетельствуя скорее об обратном ходе событий.
В этом конкретном примере решение достаточно очевидно: человеческий череп вторично приобрел эмбриональные черты в результате относительного замедления соответствующих онтогенетических процессов. В других же случаях такой ясности может и не быть. Представляет ли собой палеонтологическая последовательность тот самый серьезный контраргумент, который делает неприемлемыми рекомендации биогенетической презумпции, или наоборот, онтогенетические данные не позволяют следовать палеонтологической презумпции - чтобы решить это, нужен опыт, интуиция и вообще искусство.
Конечно, и здесь на помощь приходят презумпции. Например, при наличии нескольких независимых источников филогенетической информации, дающих противоречивые указания, нужно следовать указаниям большинства. Действительно, в полном согласии с палеонтологической презумпцией целый ряд других человеческих органов (прежде всего мозг, но также вывернутые губы, резко дифференцированный волосяной покров и т.д.) имеет достаточно выраженный продвинутый (апоморфный) характер. Также и первичная структура ДНК тесно сближает человека не с человекообразными в целом, а именно с шимпанзе. Это опять-таки противоречит биогенетической презумпции, которая предполагает близость человека к корням всех человекообразных, а не одного шимпанзе. Однако мнение большинства тоже не всегда глас божий: когда в меньшинстве оказываются источники информации, зарекомендовавших себя как особо надежные, нужно следовать им вопреки советам большинству. Если же мы станем разбираться дальше, то найдем еще массу оговорок.
Как легко видеть, использование презумпций не есть простое дело, оно всегда требует опыта, интуиции и смелости, поскольку риск ошибки никогда не бывает пренебрежимо мал. Но такова жизнь, а наука тем более. И другого пути познания я себе не представляю.
МЕТОДОЛОГИЯ ФИЛОГЕНЕТИКИ
Филогения понимается как история конкретной группы в терминах уменьшающегося генеалогического родства и изменяющегося сходства между организмами. Эволюция обычно протекает слишком медленно, чтобы ее можно было наблюдать непосредственно, так что филогения познаваема только через ее результаты. Эти результаты - признаки формы и функции организмов, включая морфологические, физиологические и поведенческие признаки, отношения организмов к биотическим и абиотическим факторам, их распространение в пространстве и во времени, и т. д.
В учебниках и публикациях нетрудно найти простые схемы и однозначные критерии родства. Однако простые правила в нашем деле слишком часто подводят, и вместо них приходится использовать более гибкий и трудный в обращении инструмент - презумпцию (Табл. 2).
Таблица 2. Презумпции филогенетики
|
область применения |
название |
формулировка |
|
общая |
познаваемости филогенеза |
любое сходство следует считать унаследованным, пока и поскольку не доказано обратное |
|
анализ групп |
палеонтологи-ческая |
из двух предположительно близко родственных групп та, что впервые появляется в более древних отложениях, признается за предковую, пока и поскольку не представлено надежных свидетельств обратного |
|
анализ различий |
палеонтологи-ческая |
трансформационную серию следует поляризовать в соответствии с палеонтологической последовательностью состояний признака, пока и поскольку не представлено убедительных свидетельств противного |
|
анализ различий |
биогенетическая |
трансформационную серию следует поляризовать в соответствии с онтогенетической последовательностью состояний признака, пока и поскольку не представлено убедительных свидетельств противного |
|
анализ различий |
аналогии |
если трансформационная серия успешно поляризована в одной группе, направление преобразований в другой группе следует считать сходным, пока и поскольку не представлено убедительных свидетельств противного |
|
анализ различий |
функциональ-ного совершенства |
из двух состояний признака то, что соответствует более эффективной адаптации, следует считать апоморфным, пока и поскольку нет убедительных свидетельств противного |
|
анализ различий |
сложности |
сложную структуру следует считать апоморфной относительно более простой, пока и поскольку нет убедительных резонов считать иначе |
|
анализ различий |
рудиментов |
состояние, обнаруживающее следы преобразования из другого состояния, следует считать апоморфным относительно последнего простой, пока и поскольку нет убедительных резонов считать иначе |
|
анализ различий |
необратимости |
если мы видим некоторое направление изменений определенно реализованным, то предпочтение следует той из конкурирующих кладограмм,которая не предполагает изменений в противоположном направлении, пока и поскольку нет надежных контраргументов |
|
анализ различий |
сохраненного распространения признака |
состояние признака, найденные только внутри группы, должно считаться апоморфным по отношению к тому, что распространено и за ее пределами, пока и поскольку нет убедительных контраргументов |
|
анализ сходств |
парсимонии |
наибольшее доверие заслуживает та клодограмма, что постулирует наименьшее число гомоплазий, - если нет серьезных оснований считать наиболее обоснованной другую кладограмму |
|
анализ сходств |
взвешенного сходства |
в случае конфликта сходств унаследованными следует считать те из них, которые по опыту изучения других групп, особенно близкородственных, известны как более надежно отражающие родство, - если нет серьезных оснований считать иначе |
Первая и центральная презумпция филогенетики это презумпция ее познаваемости: любое сходство следует считать унаследованным, пока и поскольку не доказано обратное. Генниг (Hennig, 1966) назвал ее "дополнительным (auxiliary) принципом", но на мой взгляд это центральное обобщение в филогенетике, утверждающее ее познаваемость. Действительно, "if homoplasy is universal, the characters imply nothing about the genealogy" ["если гомоплазия (независимо приобретенное сходство) универсальна, признаки не несут никакой информации о генеалогии"] (Farris, 1983: 14).
Здесь не все так уж очевидно. Человек всегда рождает человека, а не обезьяну, независимо от условий и обстоятельств, и здесь сходство однозначно указывает на происхождение. Напротив, вода остается водой, получена ли она сжиганием водорода или окислением липидов в организме. Дело здесь в разной сложности объектов, из-за которой живые организмы меняются в тесной зависимости от их истории, а превращения химикалий существенно контролируются обстоятельствами и простыми правилами, но не историей. Но живые существа также различны по уровню сложности. Как уже упоминалось, у прокариот объем независимо приобретенного сходства столь велик, что традиционными методами расшифровать их филогению не удается (Заварзин, 1987). Такие наблюдения с необходимостью ограничивают возможности изучения филогении.
Существуют и не столь экзотические ограничения. Расхождение низших таксонов происходило в не столь отдаленные времена, как дивергенция крупных групп, и следы этих событий должны была бы сохраниться лучше. Но генеалогические отношения между видами, особенно в крупных родах, обычно менее ясны, чем между семействами. Я вижу только одно приемлемое объяснение этого феномена: гомоплазия, т.е независимо возникшее сходство, вещь гораздо более обычная, чем мы привыкли думать, и она особенно обычна у близко родственных форм, поскольку их сходная организация предопределяет и сходную реакцию на сходные изменения.
Действительно, когда группа тщательно изучается и на палеонтологическом, и на современном материале, обилие гомоплазий и широкое распространение параллельной эволюции становится особенно наглядным, в том числе и на высших таксономических уровнях. Так позвоночные демонстрируют бесчисленные примеры многократного и независимого достижения определенных уровней организации - уровней амфибий, рептилий, млекопитающих и птиц. (Татаринов, 1976, 1987; Шишкин, 1987; Воробьева, 1992). Под старым термином аристогенез (Osborn, 1934) эта закономерность обсуждается Раутианом (1988).
Параллельная эволюция в близко родственных группах обычно описывается как проявление определенной эволюционной тенденции, свойственной группе. Эта тенденция может быть понята как эпифеномен базовой синапоморфии (underlying synapomorphy: Saether, 1979, 1986; термин сходный, но не идентичный ключевому ароморфозу Иорданского, 1977, 1979). Понятие базовой апоморфии подразумевает апоморфию (любое изменение в структуре или функции), которая была приобретена общим предком группы и которая индуцирует сходные изменения у некоторых или многих членов группы. Например, приобретение упругого продольного стержня (хорды и наследующего ей позвоночника), позволившее аккумулировать энергию изгибания тела, позволило различным хордовым независимо приобретать рыбообразную форму при переходе к активному плаванию (киты, сирены, ихтиозавры, головастики и т.д.). Другой пример, реберное дыхание как отдаленная причина перехода к гомойотермии, обсуждается Иорданским в упомянутых публикациях. В отличие от этих примеров, мы редко знаем непосредственные причины, ответственные за независимое приобретение определенных признаков членами определенной группы организмов. Тем не менее я думаю, что в основе большинства случаев массовой параллельной эволюции лежит реальная общая причина - некая однажды приобретенная особенность, т.е. апоморфия (отсюда и название underlying synapomorphy).
Широкое распространение параллельной эволюции не позволяет нам относиться к таксону как к простому продукту дивергенции: свой вклад должна вносить и параллельная адаптация близкородственных форм к сходным нишам, особенно в комбинации с процессом редукции архаического разнообразия (термин Мамкаева, 1968). Этот процесс может быть описан как созревание таксона, т.е. накопление сходных изменений в сочетании с элиминацией несбалансированных (инадаптивных) промежуточных групп. Этот подход возвращает нас к старой и хорошо известной гипотезе Дарвина о гиатусе, разделяющем таксоны, как в какой-то (возможно, в большой) мере продукте вымирания промежуточных форм. Созревание таксона может занимать много времени, отчего старшие таксоны выглядят более четко разграниченными и допускают более определенное толкование из родственных связей, чем младшие.
Феномен созревания таксона создает проблемы для реконструкции филогенеза. Мы способны восстановить общую форму филогенетического древа, но без большой уверенности в деталях, так что обсуждать вопрос о предковом виде, скажем, для семейства обычно не имеет смысла. Нередко детальное и разностороннее исследование не выявляет даже отрядную принадлежность предка исследуемого класса - при том, что реальный предок вполне может быть давно найден и описан, но не идентифицирован как предок (Воробьева, 1992). При недостатке данных древо демонстрирует обилие неразрешенных (unresolved) множественных развилков вместо простых дихотомий, которые при более детальном анализе могут превратиться в развилки, уходящие далеко вглубь предкового таксона, а таксон-потомок оказывается обладателем нескольких корней вместо одного. Впрочем, проблема полифилии - это уже из области таксономии, а не филогенетики, и подробнее о ней речь ниже.
Остальные филогенетические презумпции могут быть объединены в два больших класса в соответствии с тем, трактуют ли они историю групп или признаков.
Анализ групп
В анализе групп главный метод палеонтологический (недавно переопределенный как стратофенетика: Gingerich, 1979, 1990). Палеонтологическая презумпция для групп формулируется следующим образом: из двух предположительно близко родственных групп та, что впервые появляется в более древних отложениях, признается за предковую, пока и поскольку не представлено надежных свидетельств обратного.
Палеонтологический метод не универсален из-за хорошо известной неполноты геологической летописи, но эта неполнота не является ни подавляющей, ни случайной. Она достаточно упорядочена, так что тафономия может предсказать, какие организмы имеют высокие шансы сохраниться в летописи, а какие нет, и помогает нам решить, когда отсутствие некоей группы в конкретном разрезе действительно указывает на ее отсутствие в соответствующем древнем сообществе, а когда такого вывода делать нельзя (Ефремов, 1950; Жерихин, 1980; Янин, 1983; Zherikhin, 2002b).
Например, самые примитивные насекомые щетинохвостки появляются в палеонтологической летописи позже, чем крылатые насекомые (соответственно в вестфальское и намюрское время каменноугольного периода; более древние находки, приписываемые щетинохвосткам (Shear et al., 1984; Labandeira et al., 1988) основаны на слишком фрагментарных остатках), но это не служит надежным свидетельством против происхождения крылатых от щетинохвосток. Дело в том, что ископаемых насекомых находят преимущественно в озерных и, особенно в палеозое, в мелководных морских отложениях. Поэтому большинство ископаемых насекомых это либо летающие, либо водные или, реже, околоводные, прибрежные формы. Щетинохвосткам это обычно несвойственно, и они захораниваются реже (Zherikhin, 2002b).
В отличие от них перепончатокрылые насекомые хорошо летают, и отсутствие ископаемых перепончатокрылых до середины триаса следует учитывать в любом анализе, использующем палеонтологическую презумпцию для групп. Другими словами, пока нет серьезных свидетельств противного, не следует искать предков перепончатокрылых среди групп, вымерших до триаса.
Эти примеры показывают, что использование палеонтологической презумпции для групп не является простым делом. Оно требует знаний и опыта в палеонтологии и смежных дисциплинах, и даже при этом ошибки неизбежны. Впрочем, таковы все пути познания, и здесь есть лишь две возможности: либо избегать ошибок, либо исправлять их. Первый путь предполагает использование лишь самых безопасных (безошибочных) методов исследования. Этим мы резко ограничиваем их арсенал, но снижаем требования к знаниям и способностям исследователя. Это путь массовой, стандартизированной науки, характерный для Запада (например, кладизм). Мне же кажется предпочтительной свобода выбора методов исследования, позволяющая делать и исправлять ошибки, даже порой оборачивая их на благо науки как источник опыта и открытий. Этот подход дает возможность каждому, и рядовому, и выдающемуся исследователю, максимально использовать свои знания и искусство.
Кроме палеонтологического, существуют другие филогенетические методы анализа групп. Прежде всего, это биогеографический метод, направленный на восстановление истории групп в терминах их географического распространения. Действительно, расселяясь, группы эволюционируют, так что история расселения должна в какой-то мере коррелировать с генеалогией, и филогенетика может извлечь из этого пользу. Именно этот подход использует кладистическая, или викариантная, биогеография. Здесь, однако, есть серьезные проблемы.
Классическая, дисперсалистская биогеография рассматривает распространение группы как результат ее индивидуального расселения через различные барьеры. В качестве основных факторов распространения здесь выступают (1) географическая структура пространства, (2) дисперсионный потенциал группы, и (3) случайности. Сходный потенциал двух групп предполагает сходство их истории, включая генеалогию, в меру сходства других факторов. Однако все они вместе редко бывают достаточно сходными, чтобы в рамках классической биогеографии можно было бы рассчитывать на хорошую корреляцию между распространением и генеалогией.
Напротив, кладистическая биогеография (ее анализ здесь основан на Humphries, Parenti, 1986) рассматривает и распространение, и филогению групп растений и животных как результат воздействия барьеров (в основном создаваемых дрейфом континентов, но также и климатическими факторами) на их некогда непрерывное распространение. Этот подход игнорирует уникальные случаи и опирается лишь на картины распространения, повторяющиеся в разных группах организмов. Такие совместимые ареал-кладограммы (кладограммы, где названия групп заменены названиями занимаемых ими территорий или акваторий) интерпретируются как свидетельство соответствующей географически зависимой сходной филогении этих групп организмов.
Кладистическая биогеография выглядит идеально соответствующей целям филогенетического анализа групп, так как повторяющиеся ареал-кладограммы позволяют прямо преобразовывать их в гипотетические кладограммы для соответствующих групп организмов. Дело, однако, не так просто. Расселение организмов, определяющееся системой барьеров и расселительным потенциалом организмов (изменчивым, но не беспорядочно), не настолько хаотично, чтобы создавать только уникальные ареал-кладограммы, отсеиваемые методами кладистической биогеографии. В тех случаях, когда ареал-кладограммы повторяют друг друга, они будут уверенно, но ложно интерпретироваться этими методами. До тех пор, пока целенаправленные исследования на подтвердят подчиненной роли расселения в формировании ареал-кладограмм, я бы не рекомендовал пользоваться биогеографическими методами реконструкции генеалогии.
Существует и более серьезная причина такой осторожности. Как классическая (дисперсалистская), так и кладистическая биогеография уделяют совершенно недостаточно внимания вымиранию, явлению не только более чем обычному, но и часто географически упорядоченному. Под влиянием этого фактора ареал-кладограммы дают картины, ошибочно интерпретируемые кладистически. Таковы, например, многочисленные кладистические реконструкции, основанные на "гондванском" распространении (обзор см. Humphries & Parenti, 1986): в большинстве своем они, вероятно, ошибочны, потому что геологическая летопись там, где она существует, обычно фиксирует былое присутствие ныне "гондванских" групп на северных материках, где они в свое время вымерли (подробнее см. Eskov, 1987, 1992).
Палеонтология южных материков во многих случаях недостаточно изучена, но все же известны и обратные ситуации, когда ныне сугубо северная группа палеонтологически фиксируется на юге (l.c.). Равным образом вымирание может быть ответственно и за амфитропические (биполярные) ареалы многих таксонов, представляющие, вероятно, не наследие гипотетической Пацифиды (Humphries & Parenti, 1986), а следы былого всесветного распространения (Разумовский, 1971; Жерихин, 1978, 1993; Eskov, 2002). К сожалению, тропическая палеонтология развита еще слабее, чем южная. Тем не менее представляется значимым отсутствие в позднем мелу надежных указаний экваториальной неморской биоты, сравнимой с современной тропической (Мейен, 1987б). Единственный имеющий отношение к делу сравнительно богатый комплекс насекомых (около 1200 экземпляров из позднемеловых (сеноманских) янтарей Бирмы, Rasnitsyn & Ross, 2000; Shedrinsky et al., 2001) не обнаруживает характерных тропических форм, но включает группы, очень характерные для позднего мела Северной Сибири и Канады (перепончатокрылые Serphites Brues и Palaeomymar Meunier; последний род доживает доныне и имеет преимущественно внетропический биполярный ареал). Представлены и другие группы с ныне северным (верблюдки, Raphidiida) и биполярным внетропическим ареалом. Среди последних интересна диплопода Phryssonotus, ныне известная из Средиземноморья, Ю. Африки, Ю. Австралии, Уругвая, Чили и Кубы (Condé, 1954; кубинская находка зонально неоднозначна, т.к. кубинские возвышенности имеют субтропический климат). Напротив, в позднеэоценовое время Phryssonotus, как свидетельствует находка в балтийском янтаре, проникал далеко в северную внетропическую зону.
Есть и более сильные свидетельства кардинального отличия былого янтареносного леса Бирмы от тропических лесов, как мы их знаем. В отличие от тропических комплексов среднетретичных мексиканского и доминиканского янтарей, равно как и от миоценовых янтарей Сицилии и Северных Апеннин (Жерихин, 1993), бирманский комплекс лишен термитов семейства Termitidae, общественных пчел и высоко социальных муравьев. Их отсутствие высоко значимо, так как эти три группы насекомых играют ключевую роль в функционировании тропических дождевых лесов (Жерихин, 1978, 1993). В частности, термиты, перераспределяя органику и обеспечивая аэрацию глубоко по всему почвенному профилю, делают возможным глубокое заякоривание высоких тропических деревьев и тем самым их прочную фиксацию в почве. Муравьи предотвращают вспышки листогрызущих насекомых, которые могли бы необратимо прерывать действие листового насоса и тем ограничивать рост деревьев в высоту возможностями одного корневого насоса, как это имеет место в умеренных широтах. Наконец, обилие социальных пчел в тропиках гарантирует там успешное опыление под сомкнутым пологом леса, куда другие опылители проникают плохо. Других функциональных аналогов этих трех групп насекомых мы не знаем ни в прошлом, ни сейчас.
Итак, можно заключить, что филогенетическое значение повторяющихся ареал-кладограмм сомнительно, если только они не подкреплены палеонтологически. Предстоит еще долгий путь к созданию работоспособной биогеографической презумпции для групп. То же, по-видимому, относится и к другим аналогичным методам, например, паразитологическому, предполагающему определенное соответствие генеалогий хозяина и его специализированного паразита. Будь этот метод достаточно надежен, он позволил бы восстанавливать филогению хуже изученного члена такой пары по аналогии с лучше изученным.
Анализ признаков
Перейдем от анализа групп к анализу признаков. Его назначение двоякое: поляризация морфоклины, т.е. определение направления эволюции признака, и проверка сходства на предмет унаследовано ли оно или возникло независимо. Первое направление обозначим как анализ различий, второе - как анализ сходств.
Анализ различий
Существует несколько методов анализа различий, т.е. определения направления эволюции признаков. Прежде всего, это палеонтологический метод, сходный с палеонтологическим методом для групп (стратофенетика, см. выше), но все же независимый от него. Гомоплазия, или независимо приобретенное сходство, не только обычное явление, но и нередко возникает в разных группах более или менее одновременно (или в разное время, но в той же последовательности) и из сходных предшественников. Таковы процессы маммализации тероморфных рептилий, ангиоспермизации голосеменных и артроподизации червей (Татаринов, 1976; Tatarinov, 1985; Krassilov, 1991; Fryer, 1996; Пономаренко, 2001). Поэтому во многих случаях мы можем использовать презумпцию, утверждающую, что трансформационную серию следует поляризовать в соответствии с палеонтологической последовательностью состояний признака, пока и поскольку не представлено убедительных свидетельств противного. Эта презумпция особенно полезна при изучении групп с богатой палеонтологией и бедной (или особо склонной к гомоплазиям) морфологией.
Другой и очень сходный метод анализа различий - онтогенетический метод поляризации морфоклин, основанный на биогенетической презумпции: трансформационную серию следует поляризовать в соответствии с онтогенетической последовательностью состояний признака, пока и поскольку не представлено убедительных свидетельств противного. Фактически это просто пересказ знаменитого "закона" Бэра (Baer, 1828, с. 224): "Aus dem Allgemeinsten der Formverhältnisse bildet sich das weniger allgemeine und so fort, bis endlich das Specialliste auftritt" ("От наиболее общих признаков [т.е. синапоморфий самого старшего таксона, - А.Р.] развитие идет к более частным свойствам [синапоморфиям подчиненных таксонов] и так далее вплоть до наиболее специальных" [синапоморфий конечного таксона]).
Распространено мнение об экстраординарной важности "биогенетического закона" как "прямого аргумента" в филогенетике (напр. Nelson, 1978; Bonde, 1984), но я бы не согласился с этим. Онтогенез гибок, и любая его стадия может быть изменена или утрачена в результате педоморфоза, преимагинальной адаптации (ценогенеза) или рационализации (спрямления) путей развития. В результате биогенетический метод высоко эффективен в изучении филогенеза одних групп и вводит в заблуждение в других: он не имеет априорных преимуществ перед другими методами и действует не как твердое правило, а лишь как презумпция. Последовательность стадий развития - это результат прямого наблюдения, но таковы же и стратиграфическая последовательность ископаемых, и их признаки, и общее распределение признаков в системе организмов. Каждый из этих трех наборов данных сохраняет какие-то следы происхождения и в меру этого отражает филогенез и, следовательно, может быть использован для его реконструкции соответственно их возможностям в каждом конкретном случае, а не согласно каким-то априорным принципам. Назидателен пример человека (Bonde, 1984): мы оцениваем строение его черепа как педоморфное только потому, что знаем топологию кладограммы приматов из других источников (прежде всего это распределение различных признаков, включая структуру ДНК). Иначе, опираясь лишь на онтогенез, признаки человеческого черепа пришлось бы признать симплезиоморфиями с гиббоном и низшими обезьянами, противопоставленными синапоморфиям крупных человекообразных обезьян.
Третий важнейший метод анализа различий - метод аналогии. Его презумпция звучит следующим образом: если трансформационная серия успешно поляризована в одной группе, направление преобразований в другой группе следует считать сходным, пока и поскольку не представлено убедительных свидетельств противного. Действительно, если мы уверены, что крыло летучей мыши - это преобразованная нога, вполне законно предположить то же и для крыла птерозавра и птицы. Сходное предположение в отношении крыла насекомого не столь очевидно из-за глубоких различий как между самими группами, так и между их крыльями. Однако и оно не вполне абсурдно, так как не исключено участие элементов ноги в формировании крыла насекомого (Kukalová -Peck, 1978; Rasnitsyn, 1981). Равным образом мы знаем, что большинство млекопитающих эволюционировало в направлении увеличении размеров тела ("закон филогенетического роста", Cope, 1904). Поэтому мы можем предполагать сходное направление эволюции млекопитающих везде, где это предположение не встречает серьезных контраргументов. То же справедливо и в отношении других так называемых эмпирических закономерностей эволюции.
Еще один хорошо известный метод функционального совершенства может быть формализован как основанный на презумпции: из двух состояний признака то, что обеспечивает более эффективное функционирование, следует считать апоморфным, пока и поскольку нет убедительных свидетельств противного. Например, живорождение считается продвинутым (апоморфным) по сравнению с откладкой яиц, поскольку оно обеспечивает лучшее выживание потомства (позволяет живородящим формам выживать несмотря на меньшую плодовитость).
Предыдущей презумпции подчинена презумпция сложности: более сложная структура требует от организма дополнительных расходов и поэтому возникает лишь в силу ее большей эффективности. Поэтому сложную структуру следует считать апоморфной относительно более простой, пока и поскольку нет убедительных резонов считать иначе. Такие резоны появляются не так уж редко, поскольку сложные структуры сами склонны к дальнейшему упрощению. Сравните примитивную лапу млекопитающего с лошадиным копытом.
Однако такие вторично упрощенные структуры часто демонстрируют рудименты или иные следы утраченной сложности. На этом основан метод рудиментов и его презумпция: состояние, обнаруживающее следы преобразования из другого состояния, следует считать апоморфным относительно последнего простой, пока и поскольку нет убедительных резонов считать иначе (например, пока не обнаружилось, что это не рудимент, а зачаток).
Можно еще вспомнить знаменитый закон необратимости эволюции, который тоже более чем не безошибочен и представляет собой просто еще одну презумпцию: если мы видим некоторое направление изменений определенно реализованным, то предпочтение следует той из конкурирующих кладограмм, которая не предполагает противоположного направления изменений, пока и поскольку нет надежных контраргументов.
Последний метод анализа различий это анализ распространения признаков. Его основанием служит презумпция сохраненного распространения признаков: то состояние признака, что найдено только внутри группы, должно считаться апоморфным по отношению к тому, которое распространено и внутри группы, и за ее пределами, - если только нет убедительных контраргументов. Анализ распространения признаков - это вероятностный тест, основанный на том, что признак, приобретенный более отдаленным предком, имеет шанс сохраниться у более широкого круга потомков, чем тот, что возник позже в эволюции той же группы.
Когда мы видим некое состояние признака представленным как внутри данной группы, так и за ее пределами, а другое состояние свойственным только этой группе, мы предполагаем (в отсутствие серьезных контраргументов), что первое состояние плезиоморфно относительно второго. Действительно, действуя таким образом, мы можем обойтись предположением о единственном событии, сопровождавшем становление нашей группы. В противном случае, принимая более широко распространенный признак как апоморфный, мы явно или неявно предполагаем как минимум два идентичных события - независимое возникновение апоморфного признака внутри и вне нашей группы. Если мы согласны, что унаследованное сходство более вероятно (более обычно), чем приобретенное независимо, то при прочих равных вторую из этих гипотез следует считать менее вероятной (менее парсимоничной: см. ниже). Так, роговая чешуя есть у всех амниот, волосы только у млекопитающих (и некоторых близких к ним рептилий, Татаринов, 1976; гомология волос птерозавров, описанных Шаровым, 1971, еще ждет своего подтверждения), а перья только у птиц (и опять-таки у некоторых родственных им рептилий, Курзанов, 1987). Эти случаи - типичные клиенты презумпции сохраненного распространения признаков: применяя ее, мы делаем предположение, что волосы и перья развились из чешуй (апоморфны по отношению к ним), а не наоборот.
В кладизме ("ручном", не компьютерном) анализ распространения признаков под названием метода внешней группы считается важнейшим, если не единственно законным (см., например, Клюге, 2000); в других случаях, правда, абсолютизируется онтогенетический метод (см. выше). В компьютерном ("экономном") кладизме, где проблема поляризации трансформационной серии странным образом вообще устранена, под методом внешней группы понимается совсем иная процедура. Подробнее об этом чуть позже.
Анализ сходства
Перейдем к анализу сходства. Как я уже говорил, он нужен, чтобы отличить унаследованное сходство от независимо приобретенного (гомоплазии). Если мы принимаем нашу первую презумпцию ("сходство признается унаследованным, пока и поскольку..."), и если это "пока и поскольку..." не имеет места, такой анализ просто не нужен. Фактически анализ сходств и производится, только когда сходство по разным признакам распределено противоречивым образом и не может быть унаследованным во всех случаях одновременно. Например, если сходство китов с млекопитающими мы признаем унаследованным, то их сходство с рыбами по плавательным адаптациям приходится считать независимо приобретенным (т.е. гомоплазией), и наоборот.
Существует два основных метода выявления гомоплазии. Один из них - метод парсимонии. Он представляет собой прямое приложение первой презумпции: если сходство в норме унаследовано и лишь более редко возникает как гомоплазия, то гомоплазию не следует постулировать без нужды (Farris, 1983: 8), и заслуживающей наибольшее доверие клодограммой является та, что постулирует наименьшее число гомоплазий, - если, конечно, нет серьезных оснований считать наиболее обоснованной другую кладограмму. Я принимаю здесь традиционную интерпретацию парсимонии, восходящую к бритве Оккама и близкую к понятиям экономии, т.е. достижения цели минимально необходимыми для этого средствами. Другое, широкое понимание парсимонии (Farris, 1983), включающее взвешивание филогенетических доказательств, делает понятие парсимонии просто синонимом наилучшей гипотезы и потому излишним.
Но почему же опять "если нет серьезных оснований..."? Почему парсимония это только презумпция, а не твердое правило, как считают многие? Просто потому, что не все признаки равноценны. Если мы рассмотрим сходство мимикрирующей бабочки с моделью, подсчитаем все сходные элементы в их окраске и сравним это число с числом сходств каждого из этих видов с их ближайшими родичами, я не уверен, что баланс сходств будет всегда в пользу реального родства. Сходство по разным признакам отражает родство с разной степенью надежности. Сходство в типе симметрии и в общей структуре основных органов мы обычно считаем филогенетически более важным, чем в окраске или в деталях структуры поверхности тела, хотя есть и исключения. Например, отсутствие синего пигмента в волосах есть признак всего класса млекопитающих, а различия в составе пигментов в систематике прокариот и растений используются как характеристики самых старших таксонов. Но в целом окраске приписывается подчиненное значение в филогенетике.
Таким образом, филогенетическое значение сходства сильно зависит от того, какие группы сравниваются и какие их признаки сходны. Поэтому взвешивание признаков при выявлении гомоплазии представляется важным. Твердые правила здесь невозможны, по крайней мере в нынешних условиях, поэтому речь идет об очередной презумпции или, скорее, о наборе презумпций, уже частично обсуждавшемся (Расницын, 1988в; Расницын, Длусский, 1988), хотя здесь требуется еще много усилий для достижения приемлемой полноты. В обобщенном виде презумпцию взвешенного сходства можно сформулировать следующим образом: в случае конфликта сходств (когда все они не могут быть одновременно признаны унаследованными) те из них следует считать унаследованными, которые по опыту изучения других групп, особенно близкородственных, уже известны как более надежно отражающие родство, - если только нет серьезных оснований считать иначе.
Пример
Продемонстрирую применение презумпций на примере филогении перепончатокрылых насекомых и в частности проблемы положения семейства Xyelidae. Это сложный случай, породивший долгие дебаты. Если не сказано иначе, использованы данные автора (Расницын, 1969, 1980, 1983б, 1988г, 1990б, 1993; Rasnitsyn, 1988) и цитированных там источников.

Рис. 3. Triassoxyela foveolata Rasnitsyn, 1964, представитель Xyelidae из вымершего триасового подсемейства Archexyelinae; средний или поздний триас Киргизии (по Расницыну, 1969).
Перепончатокрылые известны со второй половины триаса, где найдены в Средней Азии и, по одному виду, в Австралии и Южной Африке. Все они принадлежат семейству Xyelidae (Рис. 3), как это подтверждается характерным строением антенн и крыльев. Это семейство (Рис. 4) существует доныне в северном полушарии в качестве реликтовой группы, объединяющей 5 родов и около полусотни видов, но до середины мелового периода это была ведущая группа перепончатокрылых, по крайней мере в условиях более умеренного климата.
Другие перепончатокрылые появляются позже и не известны до ранней юры (Рис. 5), что позволяет применить палеонтологическую презумпцию для групп и предложить ксиелид на роль предков остальных перепончатокрылых. Эта гипотеза согласуется с большинством других наблюдений, представляющих семейство как наиболее плезиоморфное в отряде по всем признакам, кроме одного, обсуждаемого далее. Действительно, жилкование крыльев ксиелид самое богатое и наиболее сходное с жилкованием других насекомых: переднее крыло с развилком на RS, как у различных более примитивных насекомых, и с дополнительными развилками на SC у некоторых вымерших видов, а заднее крыло - со свободным основанием Cu. То же справедливо для строения тела и предполагаемого исходного образа жизни. В частности, существенно в этой связи предположение о развитии исходных ксиелид внутри мужских шишек голосеменных, возможно, унаследованное от древнейших насекомых с полным превращением (Рис. 6). Однако плезиоморфии кладистически несущественны, поэтому рассмотрим подробнее строение антенны ксиелид, единственный признак, отвергнуть апоморфный характер которого не так просто. Что касается подразделенной галеи (лопасти ротового аппарата), то этот апоморфный признак может быть более позднего, послетриасового происхождения: он известен только в ветви, образованной более молодыми подсемействами Xyelinae и Macroxyelinae, но не известен для триасовых Archexyelinae.

Рис. 4. Раннемеловые Xyelidae из Забайкалья: а - представитель современного рода Xyela (Mesoxyela) mesozoica Rasnitsyn, 1965 (Xyelinae) (по Rasnitsyn, 1995), б - Anthoxyela turgensis Rasnitsyn, 1990, вымерший род из второго современного подсемейства Macroxyelinae (по Расницыну, 1990б).
По сравнению с нормальной антенной насекомого ксиелидная антенна несомненно апоморфна, поскольку несколько базальных члеников ее жгутика слиты в длинный и толстый сложный "третий" членик, несущий на вершине жгутик из обычных (коротких и тонких) члеников. Апоморфная природа такой структуры подтверждается применением нескольких презумпций из числа рассматривавшихся ранее. Это относится к палеонтологической презумпции для групп, поскольку наиболее близкородственная перепончатокрылым и более древняя группа (отряд Palaeomanteida = Miomoptera, Рис. 6) имела нормальные антенны. Приложима здесь и аналогичная презумпция для признаков, так как никто из известных дотриасовых насекомых не обладал похожими антеннами. То же справедливо и для биогенетической презумпции, поскольку на ранних стадиях развития (Рис. 7) сегментация антенны более равномерна, чем у взрослого насекомого, и для презумпции аналогии: сильно увеличенный, хотя и не составной по происхождению третий членик антенны мухи (Рис. 8) несомненно апоморфен. Наконец, о том же говорит презумпция сохраненного распространения признаков: "ксиелидные" антенны известны только у перепончатокрылых, а нормальные - как в этом отряде, так и за его пределами.
Нет сомнений, что на уровне отряда ксиелидные антенны апоморфны, т.е. что они возникли не раньше, чем у общего предка перепончатокрылых. Соблазнительно распространить этот вывод и на внутриотрядные отношения, тем более что нормальные антенны у перепончатокрылых распространены гораздо шире, чем ксиелидные. Однако этот вывод, по-видимому, ошибочен. Действительно, хотя из современных насекомых такие антенны в их полном выражении известны только у ксиелид, у некоторых других низших перепончатокрылых (пилильщиков и рогохвостов) антенны могут быть описаны как различным образом модифицированные ксиелидные антенны (Рис. 9). Наименее изменены по сравнению с ними антенны в тентрединоидных семействах Blasticotomidae и Argidae, где третий членик еще сильнее гипертрофирован, чем у ксиелид, но последующие членики редуцированы до одного (некоторые бластикотомиды, Рис. 9а) или полностью (Рис. 9б).
Другой источник доказательств - антенны современных паутинных (памфилиоидных) пилильщиков и настоящих (сирикоидных) рогохвостов. У Pamphiliidae (кроме некоторых видов Pamphilius) третий членик антенны подразделен, но сравнительно мал: лишь в несколько раз длиннее четвертого и почти одной с ним толщины (Рис. 9в). Менее известна, но не менее важна антенна примитивного рогохвоста рода Brachyxiphus Philippi (Siricoidea, Xiphydriidae), обнаруживающая довольно длинный и толстый "третий" членик, приблизительно равный по длине трем последующим вместе, а у самца B. grandis Philippi к тому же дополнительно подразделенный. Большинство, если не все другие ксифидрииды также демонстрируют заметно увеличенный третий членик (Рис. 9г), так что этот признак, видимо, является исходным для семейства.

Описанное распределение сложного третьего членика антенн можно было бы объяснить его независимым развитием в 5 разных группах низших перепончатокрылых. Однако это число удваивается, если учесть данные по роду Pamphilius (Van Achterberg, Van Aartsen, 1986: tab. 1, 2, fig. 366) и не предполагать, что сложное строение третьего сегмента может быть утеряно. Еще больше увеличивается это число, если принять во внимание палеонтологический материал.

Коллекция Палеонтологического института РАН в Москве содержит подавляющее большинство юрских и раннемеловых перепончатокрылых мира, в основном происходящих из Казахстана, Сибири и Монголии (данные на 1994 г.). Из них 515 принадлежит пилильщикам и рогохвостам (отряд низших перепончатокрылых Siricina), 120 из которых позволяют установить строение антенн. Из них 65 принадлежат Xyelidae и обладают соответствующим строением антенн, а из оставшихся 55 ископаемых 23 также демонстрируют характерные ксиелидные антенны (включая те, где третий членик не столь велик, как у ксиелид, но явственно толще и намного длиннее каждого из последующих), и лишь 32 обладают антеннами, не измененными или почти не измененными в сторону "ксиелидности".
Рис. 6. Palaeomantina pentamera Rasnitsyn, 1977 (Palaeomantiscidae, отряд миомоптеры, Palaeomanteida); ранняя пермь Урала (реконструкция А.Г. Пономаренко, из Родендорфа и Расницына, 1980).
Рис. 7. Антенна куколки Xyela julii Brébisson, 1818 (Xyelidae) (Расницын, 1968). I-XII = членики антенны, "III" = сложный "третий" членик антенны.

Рис. 8. Антенна примитивной мухи Glutops semiformis Nagatomi & Saigusa, 1970 (Rhagionidae) (по Nagatomi, 1982).
Среди ископаемых, не принадлежащих ксиелидам, но обнаруживающих характерные для них антенны, можно видеть, во-первых, четыре рода (Xyelocerus Rasnitsyn, Xyelotoma Rasnitsyn, Pseudoxyela Rasnitsyn и Dahurotoma Rasnitsyn, Рис. 10а-г, соответственно), демонстрирующие характерные синапоморфии современных Tenthredinoidea: переднее крыло с 2r-rs, впадающей в RS дистальнее 2r-m, и ячейку 1mcu, увеличенную таким образом, что первый отрезок RS укорочен или утрачен вовсе. У большинства родов также ствол SC слит (правда, только дистально) со стволом R, так что дистальная ветвь SC, если развита, превращена в поперечную жилку, как у современных тентрединоидов (исключение составляет Pseudoxyela Rasnitsyn, сохраняющая свободный ствол SC и, по-видимому, образующая сестринскую группу по отношению к остальным тентрединоидам). Эти четыре рода выделены в вымершее семейство Xyelotomidae, синапоморфное с современными тентрединоидами по строению жгутика антенны, сравнительно толстого и с уменьшенным числом члеников (в предположении, что многочленистый жгутик Diprionidae и некоторых Pergidae вторичен). Xyelotomidae лишены апоморфий по отношению к другим тентрединоидам и потому занимающее положение их предка.
Следующие пять родов с антеннами того же типа, что у Xyelidae, обнаруживают характерную синапоморфию надсемейства Pamphilioidea, укороченный когтевидный яйцеклад, занимающий то же положение в теле. Общий облик и жилкование крыльев также сходны с характерными для современных памфилиоидов. Среди этих ископаемых Xyelyda Rasnitsyn (Рис. 11а), Prolyda Rasnitsyn (Рис. 11б) и Mesolyda Rasnitsyn (Рис. 11в) образуют вымершее семейство Xyelydidae, лишенное синапоморфий и, следовательно, предковое для других памфилиоидов, а Praesirex Rasnitsyn (Рис. 11г) и Turgidontes Rasnitsyn (Рис. 11д), синапоморфные с современными Megalodontidae по утрате SC, объединены в вымершее семейство Praesiricidae, предположительно предковое для Megalodontidae.

Рис. 10. Юрские и меловые пилильщики семейства Xyelotomidae: a, Xyelocerus admirandus Rasnitsyn, 1968, б, Xyelotoma nigricornis Rasnitsyn, 1968, в, Pseudoxyela heteroclita Rasnitsyn, 1968 (все из поздней юры Южного Казахстана), г, Dahurotoma robusta Rasnitsyn, 1990, ранний мел Забайкалья (а, б - по Расницыну, 1969, в - по Rasnitsyn, 1996, г - по Расницыну, 1990б).
Еще одна группа ископаемых, охарактеризованная ксиелидными антеннами, включает несколько родов (Рис. 5, 12а-д), ныне объединенных в семейство Sepulcidae (Расницын, 1993). Сепульки отнесены к надсемейству Cephoidea на основании синапоморфий с современными Cephidae (стеблевыми пилильщиками) по умеренно и равномерно узкому костальному полю (иногда вторично сильно суженному в основании) и с SC, слитой с R (иногда кроме основания) и лишенной конечной ветви, пересекающей костальное поле. Сепульки лишены характерных апоморфий стеблевых пилильщиков и вероятно были их предками.


Последняя группа с ксиелидными антеннами очерчена менее четко. Я включаю туда Protosirex Rasnitsyn (Рис. 13), который на основании внешнего облика и некоторых плезиоморфий (наличие свободной SC) был отнесен к рогохвостам (Siricoidea), но может представлять группу, предковую как для них, так и для цефоидов. Вместе с некоторыми другими юрскими ископаемыми, Protosirex отнесен к вымершему семейству Gigasiricidae. Другие рогохвосты с ксиелидными антеннами - это Syntexyela media Rasnitsyn и S. inversa Rasnitsyn (Рис. 14а-б), принадлежность которых к сирикоидам несомненна из-за сочетания характерной синапоморфии (сильный игловидный яйцеклад) и плезиоморфии (сохранение свободной ветви SC). Они представляют преимущественно мезозойское семейство Anaxyelidae (с единственным современным видом), что подтверждается синапоморфией по редукции ствола SC и поперечной 2r-m переднего крыла и 1r-m заднего. Существенно, что третий членик антенны у этих двух видов модифицирован по сравнению с антенной Xyelidae, и модифицирован по-разному: он внешне цельный, но заметно уменьшен в размерах у S. inversa, и большой, как у ксиелид, но явственно подразделен на первичные членики у S. media. У близких родов анаксиелид антенны либо внешне нормальные, либо третий членик лишь немного больше других (Рис. 14в-г; подробнее см. Расницын, 1969).


Рис. 12. Юрские и меловые пилильщики семейства Sepulcidae с ксиелидными антеннами: a, Xyelula hybrida Rasnitsyn, 1969, б, Onokhoius aculeatus Rasnitsyn, 1993, в, Parapamphilius confusus Rasnitsyn, 1968, г, Pamparaphilius mongolensis Rasnitsyn, 1993, д, Micramphilius minutus Rasnitsyn, 1993; из поздней юры Южного Казахстана (a, в) и нижнего мела Забайкалья (б, д) и Монголии (г) (по Расницыну, 1993).

Теперь мы можем оценить значение антенн для понимания ранней эволюции перепончатокрылых. Фактически мы обнаружили здесь конфликт предполагаемых синапоморфий: перечисленные выше рода предположительно синапоморфны с ксиелидами по строению их антенн, тогда как по другим признакам они предположительно синапоморфны с некоторыми другими группами пилильщиков или рогохвостов. Эти гипотезы не могут быть справедливы одновременно, т.е. упомянутые сходства не могут все быть синапоморфиями. Наша задача теперь состоит в том, чтобы оценить, какие из предполагаемых синапоморфий могут быть фальсифицированы, т.е. переквалифицированы как симплезиоморфии либо гомоплазии. Только тогда можно будет построить правдоподобную и непротиворечивую кладограмму.
Можно предложить четыре различных гипотезы:
1) сходство по наличию ксиелидной антенны синапоморфно, а альтернативные признаки сходства являются симплезиоморфиями,
2) то же, но альтернативные признаки сходства являются гомоплазиями,
3) альтернативный набор сходств представляет синапоморфии (перечисленные группы родов монофилетичны с указанными выше семействам и надсемействами, но не с ксиелидами), а ксиелидные антенны приобретены независимо (являются гомоплазиями),
4) как выше, но сходство с ксиелидами по строению антенн - симплезиоморфия (унаследовано от общего предка).


Рис. 14. Юрские рогохвосты семейства Anaxyelidae с модифицированными ксиелидными (а, б) и более или менее нормальными антеннами (в, г): a, Syntexyela media (Rasnitsyn, 1963), б, S. inversa Rasnitsyn, 1968, в, Anaxyela gracilis Martynov, 1925, г, Brachysyntexis micrura Rasnitsyn, 1969; все из поздней юры Южного Казахстана (a, в, г, по Расницыну, 1969, б, по Rasanitsyn, 1996).
Гипотезы (1) и (2) означают, что все обсуждаемые рода монофилетичны с ксиелидами (кладистически принадлежат этому семейству), а все альтернативные признаки сходства, сближающие их с другими семействами и надсемействами низших перепончатокрылых, либо унаследованы от каких-то более далеких предков (гипотеза 1), либо приобретены независимо (гипотеза 2). Гипотеза (1) наименее правдоподобна, потому что она подразумевает направление эволюции соответствующих признаков противоположным тому, которое поколения исследователей принимали как нормальное (т.е. наиболее обычное и поэтому отвечающее соответствующим презумпциям). Это представляется возможным в одних случаях, но маловероятным в других. Например, трудно поверить, что своеобразно редуцированный и измененный яйцеклад паутинных пилильщиков (Рис. 11а-б) и мощный игловидный яйцеклад рогохвостов (Рис. 14а,в,г) являются (оба!) предшественниками пиловидного яйцеклада ксиелид и тентрединоидов. Действительно, несколько презумпций свидетельствуют в пользу обратного направления изменений. У большинства не перепончатокрылых насекомых с полным превращением, включая Palaeomanteida, предполагаемых предков перепончатокрылых, яйцеклад листо- или пиловидный (Рис. 6) либо (чаще) редуцирован. Кроме перепончатокрылых, игловидный яйцеклад известен у сверчков и у вымерших равнокрылых семейства Archescytinidae: там он в обоих случаях явно апоморфен. К этим данным применимы обе палеонтологические презумпции, презумпции сохраненного распространения признаков и аналогии, и все они свидетельствуют в пользу плезиоморфного характера пиловидного яйцеклада пилильщиков. Равным образом презумпции однозначно поддерживают апоморфность редуцированного жилкования крыльев анаксиелид и плезиоморфность полного жилкования ксиелид и паутинных пилильщиков. В противоположность последующим гипотезам, я не в состоянии проиллюстрировать этот гипотетический вариант соответствующей кладограммой из-за несовместимости вытекающих из нее заключений о поляризации разных признаков, как это было видно на примере яйцеклада.
Вторая гипотеза, утверждающая гомопластическую природу сходства обсуждаемых родов с иными, чем ксиелиды, группами пилильщиков и рогохвостов, не столь противоречива. Однако она предполагает очень большой объем гомоплазий, к которым придется отнести все перечислявшиеся ранее признаки сходства, альтернативные сходству этих родов с ксиелидами (легенда к Рис. 15, курсив). Это кажется маловероятным, поскольку означало бы ошибочность результатов, полученных предшествующими поколениями систематиков, которые считали соответствующие таксоны естественными, а их признаки - не слишком склонными к многократному независимому возникновению (т.е. поддерживаемыми презумпцией взвешенного сходства).

Рис. 15. Кладограмма низших перепончатокрылых (подотряд Siricina), построенная в соответствии с гипотезой (2) (см. текст), согласно которой сходство по наличию ксиелидной антенны представляет синапоморфию, а альтернативные сходства - гомоплазию. Цифры под линиями родства соответствуют наборам синапоморфий (см. ниже) (по данным Rasnitsyn, 1988, а для Pamphiliidae, Van Achterberg & Van Aartsen, 1986); гомоплазии выделены курсивом (по Rasnitsyn, 1996, с изменениями):
Есть и еще одна причина считать обе предыдущие гипотезы маловероятными - строение антенн двух видов Syntexyela. Действительно, антенна S. media легко интерпретируется как промежуточный этап трансформации нормальной антенны в ксиелидную путем утолщения и слияния базальных члеников жгутика (либо наоборот, распадение сложного третьего членика на составляющие его первичные, но в рамках двух рассмотренных гипотез это исключенный вариант). Напротив, в антенне S. inversa и некоторых других видов того же рода третий членик цельный, но заметно увеличенный, хотя и далеко не так, как в типичных ксиелидных антеннах. Он выглядит как этап превращения ксиелидных антенн в нормальные путем уменьшения сложного третьего членика до размеров простого, что также невозможно в рамках обоих этих гипотез, либо как попытка независимого создания ксиелидных антенн путем простого увеличения первого членика жгутика, что противоречит сложному строению увеличенного членика там, где это можно проверить (у различных современных низших перепончатокрылых). В любом случае гипотезы (1) и (2) несовместимы с близким родством видов Syntexyela, и если мы принимаем любую из этих гипотез, все признаки сходства S. inversa с другими видами рода мы должны расценивать как гомоплазии.

Рис. 16. Кладограмма низших перепончатокрылых (подотряд Siricina), построенная в соответствии с гипотезой (3) (см. текст), согласно которой сходство по наличию ксиелидной антенны представляет гомоплазию, а альтернативные сходства - синапоморфию; в остальном как на Рис. 15 (по Rasnitsyn, 1996, с изменениями).
Обратимся теперь к альтернативным гипотезам (3) и (4). Обе они подразумевают, что сходство обсуждаемых родов (обладающих ксиелидными антеннами, но не включенных нами в Xyelidae) с соответствующими таксонами низших перепончатокрылых (обладающими нормальными антеннами) носит синапоморфный характер. Различие состоит в том, полагается ли сходство по наличию ксиелидных антенн гомоплазией или симплезиоморфией: гипотеза (3) предполагает, что сложный третий членик антенн был приобретен многократно и независимо (Рис. 16), а гипотеза (4) - что он возник единственный раз у общего предка и затем много раз был независимо утрачен (Рис. 17). Обе гипотезы кажутся более правдоподобными по сравнению с двумя первыми, предполагая всего лишь гомоплазию по единственному признаку (многократное приобретение или утрата гипертрофированного "третьего" членика антенн) и не требуют опровержения существующих представлений о путях эволюции многочисленных других признаков.
При всем том последние обсуждаемые гипотезы трудно счесть одинаково хорошо обоснованными. Гипотеза (3) многократного возникновения ксиелидных антенн требует большего числа гомоплазий, чем гипотеза (4) многократной их утраты (ср. Рис. 16 и 17, курсив), и получает поддержку со стороны презумпции парсимонии. Еще более четкий результат дает применение презумпции взвешенного сходства. Действительно, возникновение сложных структур обычно не считается простым делом, и их многократное независимое возникновение предполагается менее вероятным, чем многократная независимая утрата. Ксиелидный членик антенн представляет сложную структуру с неясной функцией, и его многократное и широко диахронное возникновение в пределах единственной компактной группы (низших перепончатокрылых) объяснить трудно. Проще предположить его однократное возникновение в отдаленные времена, в неизвестных условиях и по пока неясным причинам, и последующую независимую утрату. Это особенно ясно в случае Syntexyela: легче представить себе редукцию ксиелидной антенны разными путями (через уменьшение размеров в одном случае и распадение на первичные членики в другом), чем создание сложной структуры с неясной функцией несколькими разными способами.
Сравнение гипотез все еще не завершено: не задействована еще одна презумпция, имеющая отношение к делу. Действительно, предыдущие рассуждения кажутся достаточно простыми и ясными, особенно по поводу гипотез (1) и (2), предполагающих синапоморфию всех перепончатокрылых, обладающих ксиелидными антеннами, по этому признаку. Насколько мне известно, таких гипотез никто и не выдвигал, и мне они понадобились просто для полноты картины. Две другие гипотезы реально обсуждаются в литературе: гипотеза (3) независимого возникновения ксиелидных антенн предложена Россом (Ross, 1937), а в развернутой форме - Кенигсманом (Königsmann, 1976). Гипотеза (4) независимой утраты ксиелидных антенн развивается мной (Расницын, 1968, 1969, 1980; Rasnitsyn, 1988, 2002a) и, как она подана здесь, выглядит явно более предпочтительной. Однако позиция Росса и Кенигсмана в действительности вовсе не так слаба: она дополнительно опирается на презумпцию необратимости эволюции, поскольку восстановление ранее утраченной гомономности антенны это типичная реверсия. Тем не менее я полагаю аргументацию в пользу однократного возникновения и многократной утраты ксиелидных антенн (т.е. в пользу несоблюдения необратимости эволюции этого признака) более убедительной. В пользу этого заключения свидетельствуют результаты проведенных в последние годы компьютерных расчетов филогении перепончатокрылых (Ronquist et al., 1999: figs. 2-3; Vilhelmsen, 2000: fig. 9; Quicke et al., unpublished). Набор таксонов и признаков в этих расчетах несколько различался, и виды Pamphilius с разным строением антенн там порознь не анализировались, тем не менее во всех случаях структура кладограммы была найдена близкой друг к другу и в целом сходной с нашими кладограммами (Рис. 16, 17). При этом в соответствии с гипотезой (4) перепончатокрылые как целое оказалась охарактеризоваными, среди прочих синапоморфий, ксиелидными антеннами, - в том числе и у Вильгельмсена (Vilhelmsen, 2000), работавшего независимо от меня.
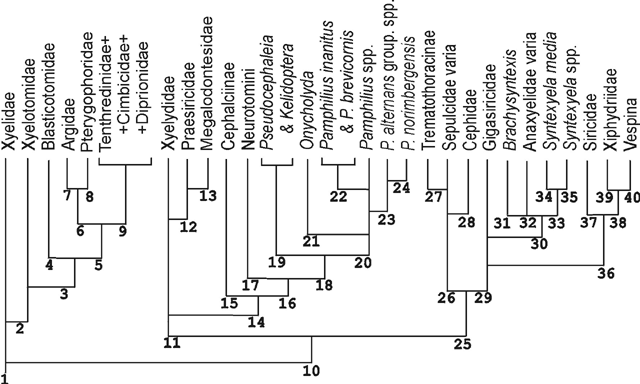
Рис. 17. Кладограмма низших перепончатокрылых (подотряд Siricina), построенная в соответствии с гипотезой (4) (см. текст), согласно которой сходство по наличию ксиелидной антенны представляет симплезиоморфию, а альтернативные сходства - синапоморфию; в остальном как на Рис. 15-16 (по Rasnitsyn, 1996, с изменениями).
Как бы то ни было, справедлива ли именно четвертая гипотеза или какая-то другая из четырех, в данном случае не самое главное. Вопрос об исходном плане строения перепончатокрылых был рассмотрен здесь не для того, чтобы установить истину, а только как пример, чтобы сделать предлагаемый подход к реконструкции кладограммы более наглядным. Хочу надеяться, что это мне удалось.
Компьютерный кладизм
Все, о чем шла речь до сих пор (кроме предпоследнего абзаца предыдущего раздела), касалось "ручной", не математизированной филогенетики. Компьютерный кладизм разработал математический аппарат, позволяющий автоматически строить кладограмму по заданному распределению признаков. Изложение этого раздела здесь изменено по сравнению с первым изданием из-за обнаруженных там неточностей. Я искренне благодарен В.А Благодерову за обнаружение и помощь в исправлении этих ошибок.
Компьютерный анализ филогении использует только две презумпции из всех рассмотренных выше: презумпции познаваемости филогенеза и презумпции парсимонии (экономии). Исходным материалом для анализа является матрица, где для каждого таксона анализируемой группы указывается состояние каждого из всех выбранных признаков (Табл. 3; на самом деле эта матрица гораздо больше и включает 236 признаков). В матрицу вводится одна или несколько "внешних групп" - таксонов, для которых предполагается положение филогенетически вне, но вблизи исследуемой группы. У Вилгельмсена это представители нескольких других отрядов насекомых. Матрицу вводят в компьютер и обрабатывают с помощью одной из специальных программ (обычно это PAUP или, теперь уже реже, Hennig86). Программа рассчитывает кладограмму самым экономным образом, так чтобы она предполагала минимум шагов (эволюционных событий: дивергенций и изменений состояний признаков). Первичным результатом компьютерных расчетов является неориентированный граф: ветвящаяся линия, свободные концы которой представляют таксоны, включенные в матрицу. Внешние группы используются для "укоренения" кладограммы, т.е. определения ее начала и, соответственно, порядка ветвления.
Таких самых коротких (экономных) деревьев обычно оказывается более одного, поэтому далее используется одна из процедур их согласования, в результате чего и появляются кладограммы с неполным разрешением - с тем или иным числом тройных и множественных развилков вместо простой дихотомии (Рис. 18). Неполное разрешение при этом понимается не как свидетельство происхождения более чем двух групп от одного предка (это в кладизме невозможно), а как результат неполноты данных, указание на то, что использованных признаков недостаточно для расчета кладограммы.
Таблица 3. Часть матрицы (первые 40 признаков) для расчета кладограммы низших перепончатокрылых (Vilhelmsen, 2001. Table 1)
0, 1, 2 -состояния признака; ? - состояние неизвестно, - - к данному таксону неприменимо.
Сокращения: Ana. - Anaxyelidae; Arg. -Argidae; Aul. -Aulacidae; Bla. - Blasticotomidae; Cep. -Cephidae; Cim. - Cimbicidae; COL. -Coleoptera; Dip. - Diprionidae; Iba. - Ibaliidae; LEP. - Lepidoptera; MEC.-Mecoptera; Meg. - Megalodontesidae; Mly. - Megalyridae; NEU. - Neuroptera; Oru. - Orussidae; Pam. - Pamphiliidae; Per. - Pergidae; PSO.-Psocodea; RAP. - Raphidioptera; Sir. - Siricidae; Ste. - Stephanidae; Ten. - Tenthredinidae; Tri.-Trigonalidae; Ves.-Vespidae; Xip.-Xiphydriidae; Xye. - Xyelidae.
Coleoptera, Lepidoptera, Mecoptera, Neuroptera, Psocodea, Raphidioptera - высшие группы.
|
Признак |
||||
|
Таксон |
1 1234567890 |
1111111112 1234567890 |
2222222223 1234567890 |
3333333334 1234567890 |
|
Amphigerontia (PSO.) |
0000001000 |
0000100-10 |
0000?110?0 |
?1-230??00 |
|
Priacma (COL.) |
0010011000 |
0??011001? |
?000?00010 |
?1-11???00 |
|
Raphidia (RAP.) |
0000000000 |
0000117001 |
?000?00000 |
01-210?100 |
|
Chrysopa (NEU.) |
0000000000 |
0?00010010 |
0000?00000 |
01-110?100 |
|
Panorpa (MEC.) |
0000-00000 |
0010010011 |
0000?01000 |
11-3200?00 |
|
Micropterix (LEP.) |
0000001100 |
0010000-10 |
0000?11010 |
1000220000 |
|
Xyela (Xye.) |
0010000100 |
0010000-00 |
0010111010 |
0001010000 |
|
Macroxyela (Xye.) |
0010000100 |
0010000-00 |
0010011010 |
01-0010000 |
|
Blasticotoma (Bla.) |
0010000010 |
0700000-?? |
?010?01000 |
?00101?010 |
|
Paremphytus (Bla.) |
0010000010 |
0??0000-00 |
002-?0100? |
?0010??010 |
|
Runaria (Bla.) |
0010000010 |
010?000-00 |
002-??1000 |
00??0?0010 |
|
Tenthredo (Ten.) |
0010000000 |
0000000-00 |
0000001000 |
?001000010 |
|
Dolerus (Ten.) |
0010000000 |
0?00000-00 |
0000001000 |
?001000010 |
|
Athalia (Ten.) |
0010000000 |
0010000-01 |
0000?01000 |
0001000010 |
|
Nematus (Ten.) |
0010000000 |
0??0000-01 |
0000001000 |
?00100?010 |
|
Monoctenus (Dip.) |
0010000000 |
0??0000-00 |
0100?0100? |
?00100?010 |
|
Gilpinia (Dip.) |
0010000000 |
0110000-00 |
0100?01000 |
0001000010 |
|
Cimbex (Cim.) |
0010100000 |
0??0001-00 |
0001001000 |
?00100?010 |
|
Zaraea (Cim.) |
0010000000 |
0010100-01 |
0001?01000 |
0001000010 |
|
Corynis (Cim.) |
0010000070 |
0??0010002 |
0001?01000 |
?0010??010 |
|
Arge (Arg.) |
0010000000 |
0000000-00 |
002-001000 |
0001000010 |
|
Sterictiphora (Arg.) |
0010000000 |
0??0000-00 |
002-?01000 |
?00100?010 |
|
Perga (Per.) |
0010000000 |
0?00001-00 |
0001?01000 |
?00210?010 |
|
Phylacteophaga (Per.) |
0010000000 |
0000000-00 |
0000001000 |
000110?010 |
|
Pamphilius (Pam.) |
0011000000 |
0??1010000 |
0000022000 |
?00100?100 |
|
Neurotoma (Pam.) |
0011000000 |
0??1010002 |
0000?2200? |
?0010??100 |
|
Cephalcia (Pam.) |
0011000000 |
0011010001 |
0000022000 |
1001000100 |
|
Acantholyda (Pam.) |
0011000000 |
0??1010000 |
0000022000 |
?0010??000 |
|
Megalodontes (Meg.) |
0011100070 |
0011010002 |
1200022000 |
000101?000 |
|
Cephus (Cep.) |
0011010020 |
0110011000 |
1000?01000 |
0001020000 |
|
Calameuta (Cep.) |
0011010020 |
0??0011000 |
1000001000 |
?00102?000 |
|
Hartigia (Cep.) |
0011010020 |
0??0011000 |
?000001000 |
?00102?000 |
|
Syntexis (Ana.) |
0011110021 |
0110011000 |
0000101000 |
0011120000 |
|
Sirex (Sir.) |
0111110020 |
0110010100 |
0000?01000 |
0003120111 |
|
Urocerus (Sir.) |
0111110020 |
0??0010100 |
0000001000 |
?00312?111 |
|
Tremex (Sir.) |
0111110020 |
0??0010100 |
0000001000 |
?00322?111 |
|
Xiphydria (Xip.) |
0111110001 |
0110110000 |
0000001000 |
0001120010 |
|
Orussus (Oru.) |
1111110001 |
1110110112 |
1000101000 |
0012101110 |
|
Schlettererius (Ste.) |
1111110001 |
1??0110102 |
1000?01101 |
?01200?010 |
|
Megalyra (Mly.) |
011---0021 |
0????1?1?? |
??00101102 |
?012101?10 |
|
Aulacus (Aul.) |
00110100?! |
0??011?102 |
10001010?? |
?01200??10 |
|
Ibalia (Iba.) |
00110?00?1 |
0??0110002 |
1000101002 |
?012101010 |
|
Orthogonalys (Tri.) |
0011170021 |
0??0110102 |
1000001101 |
?00110?110 |
|
Vespula (Ves.) |
00111100?! |
011011?112 |
1000101102 |
?011001110 |
Рис. 18. Кладограмма низших перепончатокрылых, построенная по матрице, часть которой показана в Табл. 3 (Vilhelmsen, 2001). Сокращенные названия семейств перепончатокрылых и отрядов (внешних групп) см. в Табл. 3.

Что касается проблемы полярности признаков, ее в компьютерном кладизме просто не существует: компьютер сам рассчитывает направление изменений, так чтобы получить самую короткую кладограмму, а если нам интересно, как, с точки зрения помпьютера, шла эволюция, мы можем считать направление изменений прямо с итоговой кладограммы, пользуясь встроенными в программу возможностями ("опциями").Таким образом, все наши веками накопленные знания об эволюционном процессе, о том, в каком направлении могут меняться те или иные признаки в в тех или иных условиях и обстоятельствах, оказываются никому не нужным хламом: компьютер это знает лучше нас.
Это не карикатура, это утверждают столпы компьютерного кладизма: "The most pervasive misunderstanding of outgroups and their use in cladistics is thus tied to the concept of "character polarization" and centers on the belief that we must know "primitive" and "derived" states before we attempt the analysis. This belief contrasts with the procedure described by Farris (1972), implicit in Farris' (1970) description of networks, in which character polarity is determined by the resultant topology after analyzing outgroup and ingroup taxa simultaneously" (Nixon, Carpenter, 1993: 419); "No account is taken of any a priory polarity decisions" (Kitching et al., 1998: 67).
Привлекательными сторонами экономного кладизма является возможность автоматической обработки больших массивов данных и более высокая объективность результатов. Впрочем, и здесь не все просто. Наоборот, именно в ручном кладизме число используемых признаков и анализируемых таксонов фактически не ограничено, а для компьютера такие ограничения существуют, и порой весьма чувствительные. Разница в том, что компьютер требует учитывать состояние всех отобранных для анализа признаков у всех сравниваемых таксонов, а ручной анализ допускает использование каждого признака только там, где исследователь считает его полезными. Второй подход, конечно, субъективен, но зато он учитывает хорошо известное свойство, что один и тот же признак в разных частях системы может иметь совершенно разное значение. Например, изменение числа зубов разного типа имеет первостепенное значение в систематике большинства млекопитающих, но не у китов (и некоторых других отрядов). При компьютерном анализе филогенеза млекопитающих вариабельность этого признака у китов сильно обесценит различия в зубной формуле других отрядов. Конечно, здесь ситуация очевидна, и мы можем просто объявить этот признак "неприложимым" к китообразным. Но в других случаях этот трюк удается не всегда, и в результате теряется масса филогенетически важной информации. Этот важный эффект можно назвать девальвацией признаков.
Претензии экономного кладизма на объективность ограничены еще и тем, что и очерчивание границ исследуемой группы, и поиск и выбор признаков для анализа, и выбор внешней группы - все это включает весьма значительный и неустранимый субъективный элемент. Будучи озабоченным именно объективностью, кладизм не уделяет внимания тем аспектам процедуры, которые не поддаются легкой объективизации, и редко обсуждает их методологию. Это не идет на пользу компьютерным реконструкциям.
Исходя из своего практического опыта как в ручном, так и компьютерном анализе филогенеза, могу сказать, что компьютерный анализ не имеет бесспорных преимуществ. Результаты расчетов разных авторов для одной и той же группы часто оказываются еще более противоречащими друг другу, чем при ручном анализе. Этого не видно в нашем примере, поскольку система низших перепончатокрылых сравнительно проста и неплохо изучена, но уже с высшими перепончатокрылыми положение хуже (см., например, Rasnitsyn, 2000), хотя и они - не самая трудная группа насекомых. Более того, и в пределах одного анализа результат часто оказывается очень неустойчивыми к минимальным изменениям параметров расчета. На Рис. 19 показаны 8 кладограмм, полученных именно таким образом с одной и той же матрицы: они различаются, и порой весьма существенно.
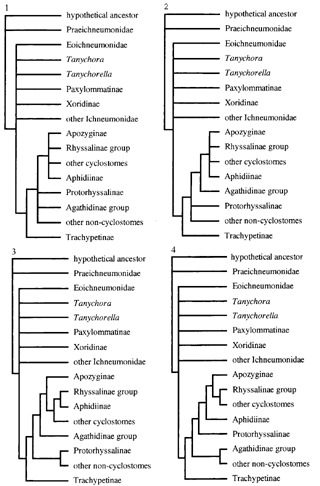
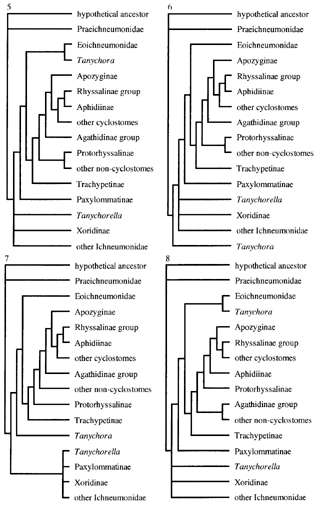
Рис. 19. Кладограммы для перепончатокрылых насекомых надсемейства Ichneumonoidea, построенные по одной и той же матрице при небольших вариациях параметров расчета (подробнее см. Quicke et al., 1999, figs. 11-12).
Самое же главное, что с углублением анализа и увеличением числа использованных признаков результат далеко не всегда улучшается - мне кажется, прежде всего из-за девальвации признаков. Этот неутешительный вывод основан прежде всего на результатах, полученных с моим собственным участием. Я имею в виду кладограммы всего отряда перепончатокрылых насекомых, рассчитанные Ронквистом и др. (Рис. 20) и Квиком с соавторами (Рис. 21), в обоих случаях при моем активном участии в отборе и анализе признаков и составлении матрицы. На мой субъективный взгляд, результат Квика не намного лучше, чем у Ронквиста, хотя его матрица включает 272 признака против 169 у Ронквиста, и вообще эта работа была задумана и выполнена как проверка и уточнение результатов Ронквиста на лучше выверенном и более широком материале. Значительного улучшения я не обнаружил, и до сих пор мои старые, весьма несовершенные результаты, полученные методом ручного кладизма (Rasnitsyn, 1988), кажется более адекватными. Не говоря уже о более поздних, которые к тому же по объективным (количественным) показателям значительно лучше согласуются с палеонтологической летописью, чем компьютерные кладограммы (Rasnitsyn, 2000, 2002а).



Рис. 20. Кладограммы для низших (а), паразитических (б) и жалоносных перепончатокрылых (в), построенные по матрице с 169 признаками (Ronquist et al., 1999, figs. 3, 8, 13). Пунктиром показаны связи вымерших семейств.
Все это я говорю не к тому, что экономный кладизм не нужен - он полезен, но не как истина в последней инстанции, а как один из подходов, результаты которого требует осмысления и проверки так же, как и результаты любого другого метода.

Рис. 21. Кладограммы для перепончатокрылых насекомых, построенные по матрице с 272 признаками (Quicke et al., не опубликовано).

Рис. 22. Кладограмма для перепончатокрылых насекомых, построенная по молекулярным данным (16S РНК, 28 S РНК и цитохромоксидаза) (Dowton, Austin, 2001, fig. 2).
Я не упоминал еще о реконструкции филогении по последовательности аминокислот в молекулах белков. Я недостаточно знаком с теорией этого метода и не буду говорить о его теоретических достоинствах и недостатках. О результатах же могу сказать, что они часто очень интересны и информативны, но в каких-то деталях часто не лезут ни в какие ворота. Это можно видеть на многих молекеулярных кладограммах: я приведу только тот пример, который мне ближе (Рис. 22). Здесь на новейшей молекулярной кладограмме перепончатокрылых типичные проктотрупоиды Heloridae оказались монофилетичны с Evaniidae, жалящие (осы, пчелы и муравьи) - с Gasteruptiidae, а примитивнейшие Orussidae попали внутрь Ichneumonidae. Никакие другие исследования даже не приближаются к этим парадоксальным результатам. Не меньшие, но другие странности обнаруживали и каждый из двух предыдущих анализов, выполненных той же группой исследователей. В первом из них (Dowton, Austin, 1994) Stephanidae попали к сирикоидам, а Orussidae - к цефоидам, во втором (Dowton et al., 1997) орехотворки оказались сестринской группой ко всем остальным высшим перепончатокрылым. В общем, молекулярный метод можно оценить как многообещающий, но пока еще он никак не может заменить другие методы.
ТАКСОНОМИЯ
Таксономическая классификация организует биологическое разнообразие таким образом, чтобы облегчить наше взаимодействие с ним. Существует много форм классификации, созданных для определенных целей и определенных категорий пользователей (например, система жизненных форм, или система вредителей по признакам причиняемого ими вреда). Они не представляют значительных теоретических проблем, и здесь речь пойдет о системе, используемой в качестве общей (междисциплинарной).
Таксоны общей системы, чтобы выполнять свои функции, должны быть осмысленными объединениями с точки зрения максимально широкого круга пользователей, как птица представляет вполне определенное понятие не только для орнитолога, но и для охотника, повара и художника. Чтобы соответствовать этой цели, таксоны общей системы должны быть максимально однородными внутри себя и максимально различными между собой по максимально широкому кругу признаков. Это свойство позволяет системе выполнять еще две важнейшие функции: свертывать информацию о живых организмах в доступной форме (вспомним, что каждый из нас знает, скажем, о воробье, включая информацию о признаках старших таксонов) и предсказывать свойства недостаточно изученных таксонов (ничтожная часть видов насекомых изучена сколько-нибудь детально, но мы не сомневаемся, что они состоят из эвкариотических клеток соответствующей структуры и функции, и с достаточной уверенностью описываем многие детали биологии даже вымерших видов).
В систематике последних десятилетий выделяются три основных направления: фенетика, кладизм и еще одно, часто неудачно именуемое "эволюционной систематикой" - в отличие от "филогенетической систематики", первоначального и столь же неудачного названия кладизма (действительно, чем отличаются термины "эволюционный" и "филогенетический" в данном контексте?). Есть для него и другие названия (синкретическая, эклектичная систематика: Farris, 1979), но это уже не названия, а скорее квалификация с точки зрения кладиста (эвфемизмы вместо "дурной систематики"). Для этого третьего направления, которое пытается выявить и сформулировать принципы традиционной систематики, недавно был предложен термин "филистика" (Rasnitsyn, 1996a).
Различия между этими направлениями гораздо глубже, чем можно было бы предположить: им соответствуют совершенно различные представления о структуре и происхождении биологического разнообразия.
Одной из этих систем взглядов принадлежит синтетическая, а более точно - популяционно-генетическая теория эволюции. Как мы видели, она представляет эволюционный процесс как динамику аллелей в популяциях, контролируемую отбором и стохастическими процессами. Отбор формирует состав популяций и свойства организмов, и делает он это с точностью, ограниченной лишь стохастическими факторами и обменом генами, который унифицирует структуру популяций на уровне, отвечающем не локальным, а усредненным характеристикам отбора.
Организация живых существ предстает здесь набором признаков, свободно тасуемых отбором, а эволюционный процесс - равномерным в той мере, в какой равномерны определяемые средой вариации отбора. Эта равномерность нарушается лишь дивергенциями и вымираниями, каковые только и могут быть использованы для построения естественной системы организмов. То есть, естественная система может быть только строго генеалогической, с границами таксонов, проводимыми по точкам дивергенции (узлам кладограммы). Таким образом, таксоны выделяются в соответствии с реконструированными фактами их истории, а не их нынешними свойствами, которые важны только как исторические свидетельства.
Это точное описание кладистической системы: строго иерархической, с не фиксированными рангами, определяемыми только последовательностью дивергенций, и с невозможностью существования парафилетических, т.е. предковых таксонов (Табл. 4).
Таблица 4. Три таксономии
|
эволюционная основа |
задающий процесс |
рабочий критерий |
сходство |
кровное родство |
|
|
Кладизм |
синтетическая |
акт дивергенции |
синапо-морфия |
игнорируется |
единственная основа системы |
|
Фенетика |
эпигенетическая |
дискретность биоразнообразия |
гиатус |
единственная основа системы |
игнорируется |
|
Филистика |
эпигенетическая |
дискретность биоразнообразия |
гиатус |
задающий критерий |
контролирующий критерий |
В отличие от синтетической, эпигенетическая теория эволюции, как мы тоже знаем, видит эволюционный процесс прежде всего как процесс эволюционного преобразования онтогенеза и в частности устоявшихся (отобранных в чреде поколений) эпигенетических процессов и их стабилизированных ансамблей - креодов. Напротив, мутации и аллели выступают здесь в роли переключателей между альтернативными траекториями развития, а не в качестве создателей структур, свойств и признаков.
Глубокая взаимозависимость между онтогенетическими процессами и, соответственно, между свойствами и признаками делает оптимизацию целого организма необычайно трудной задачей, продуктом напряженного компромисса. Однажды достигнутый успешный компромисс оказывается прочно стабилизированным (забуференным), поскольку удачная структура организма только с большим трудом поддается изменению за пределы нормальной внутривидовой изменчивости (которая сама уже стабилизирована отбором предыдущих поколений). Поэтому эволюция (успешный выход за пределы стабилизированного эпигенотипа) затруднена, мало предсказуема и более или менее скачкообразна, а возникающее в результате биоразнообразие - более или менее дискретна. Эту дискретность эксплуатируют как фенетика, так и традиционная систематика, строящие свои системы путем выявления и прослеживания гиатусов - разрывов непрерывности биоразнообразия. Но обо всем по порядку.
Кладизм
Глубинное понимание кладизма разными его последователями отличается столь сильно (ср. Hennig, 1966, и Platnick, 1979), что предлагается, например, использовать старое название "филогенетическая систематика" для версии Геннига, а кладизм оставить для так называемого трансформированного кладизма (de Queiroz & Donoghue, 1990a; это предложение не получило большой поддержки, и я сохраняю здесь широкое понимание кладизма). Однако центральное утверждение кладизма, что "фенетическая классификация строится путем группирования по общему сходству, тогда как филогенетическая - по предполагаемым синапоморфиям" (Farris, 1979: 478), остается общепризнанным. Обосновывая его, Генниг (Hennig, 1966) писал, что таксоны, обозначенные синапоморфиями, образуют систему, изоморфную филогении и потому наиболее эффективную. Отсюда и название системы - филогенетическая. Но есть и другая точка зрения, что, во-первых, само устройство (картина, pattern) природы таково, что система организмов может быть отображена ветвящейся диаграммой (иерархической классификацией), во-вторых, эта картина (pattern) может быть отображена в результате изучения признаков и поиска иерархического (internested) набора синапоморфий, и в-третьих, что наши знания об эволюции, как и о системе, выводятся из этого иерархически организованного набора (паттерна) синапоморфий (Platnick, 1979: 538). Таким образом, паттерн признается первичным, а филогения - вторичной. Такой подход ставит вопрос: кто привносит паттерн в природу? каков его генерирующий процесс, если это не эволюция? Пока на этот вопрос не предлагается ответа, я не вижу возможности отдельного анализа паттерн-кладизма (трансформированный кладизма) и возвращусь к его классической версии.
Первый возникающий вопрос касается того, какой именно смысл вкладывается в центральное положение кладизма (синапоморфия как единственное обоснование таксона). Синапоморфия обычно определяется как состояние признака, приобретенное предковым видом группы и потому свойственное всем ее членам и только им. Это определение неточно, поскольку любое приобретенное состояние признака может быть затем утрачено, что невозможно для апоморфии (змеи утратили ноги, но не четырехногость как синапоморфию тетрапод). Апоморфия - не состояние признака, а факт, что такое состояние однажды было приобретено (факт истории группы).
Специального рассмотрения заслуживает вопрос о группах, лишенных апоморфий. Такие группы именуются парафилетическими или полифилетическими в зависимости от того, выделены ли они соответственно по симплезиоморфии или гомоплазии. Кладистическое решение состоит в разбиении таких групп на меньшие таким образом, чтобы все они стали монофилетическими (точнее, голофилетическими, см. ниже), т.е. были охарактеризованы апоморфиями. При этом, однако, возникает проблема парафилетических таксонов, которые не удается расчленить на голофилетические составляющие (метатаксоны по de Queiroz & Donoghue, 1990b). Примером могут служить многие из таксонов перепончатокрылых, которые выше были охарактеризованы как предковые для других групп. В большинстве случаев эту проблему можно было бы посчитать временной и списать на неполноту знаний, которая будет преодолена. Однако это убежище неведения не спасает хотя бы потому, что параллельная эволюция, как мы видели, не оставляет нам надежды на расшифровку кладограммы во всех ее деталях.
Еще более существенно, что обязательное разделение парафилетического таксона на голофилетические подгруппы неизбежно приведет нас к предковому виду. Обычное утверждение, что предковый вид обязательно исчезает, дав начало двум дочерним видам, ни на чем не основано. Нам не известно причин, почему после превращения одной или нескольких популяций предкового вида в новый вид, остальные не могли остаться без изменений видового уровня, а затем дать начало еще нескольким новым видам (см. ниже). Таким образом, кладизм предписывает расчленить предковый вид таким образом, чтобы каждая сестринская пара могла иметь собственного предка, что сделать невозможно (de Queiroz & Donoghue, 1990a).
Некоторые исследователи и здесь прибегают к убежищу неведения, утверждая. что предковый характер вида никогда не может быть верифицирован (Nelson, 1973a). Однако, как показал Поппер, научные гипотезы в принципе не могут и не должны верифицироваться. Да и нет никаких биологических факторов и процессов, которые препятствовали бы предковому виду выжить после того, как какая-то или какие-то из его популяций превратились в другой вид, и повторить это еще и еще раз. Такие виды многократно выявлялись в более детальных палеонтологических исследованиях, особенно ярко в многочисленных работах по млекопитающим плейстоцена. То же самое прекрасно видно на обширных материалах по неогену Паратетиса, позднетретичной системы эпиконтинентальных морей к северу от остатков океана Тетис. Изменчивые гидрологический режим этих морей и система их связей обеспечили бурные процессы диверсификации и вымирания их фауны, а обилие ископаемых и экономическое значение нефтеносных отложений Паратетиса обусловили их хорошую изученность (см. Невесская и др., 1986).
В конце концов, если каждая ветвь филогенетического древа имеет в основании предковый вид, то простой расчет показывает, что таковые должны составлять половину всех когда-либо существовавших видов. Действительно, в случае строгой дихотомии ветвления и при отсутствии филетического видообразования число предковых видов равно числу терминальных видов минус единица. Множественное ветвление (почкование, полихотомия) несколько уменьшает долю предковых видов, филетическая эволюция - увеличивает их число, но в первом приближении несомненно, что половина всех описанных ископаемых видов (и, вероятно, немалое число современных) - непосредственные предки других видов.
Вид, предковый более чем для одного таксона, представляет непреодолимую теоретическую проблему для кладизма, если только не смягчить его центральное утверждение (таксон определяется исключительно через апоморфию). Этот необходимый шаг был сделан еще Геннигом, который считал, что апоморфия узаконивает таксон не по определению, а лишь как следствие из другого, более жесткого (базового) утверждения об изоморфии между системой и филогенезом. "Время существования вида определяется двумя событиями видообразования: тем, которому этот вид обязан своим возникновением как независимое репродуктивное сообщество, и тем, которое разделяет его на два или более таких сообществ" (Hennig, 1966: 66). Этот путь рассуждений делает дивергенцию единственным процессом, создающим таксоны, а апоморфия оказывается просто маркером дивергенции и, как следствие, таксона. Другим следствием будет исчезновение предкового вида в результате дивергенции - даже если один из продуктов дивергенции не обнаруживает ни малейших отличий от предкового вида (лишен апоморфий). Это утверждение выглядит несколько странным, но оно по крайней мере последовательно.
Таким образом, в кладистической парадигме некоторые (возможно, многие) виды не имеют отличий от своих предков. Тогда у нас нет даже теоретической возможности точно восстановить топологию кладограммы: возможна лишь некоторое приближение к ней. Это делает преимущества подхода не столь очевидными и открывает другие возможности, например, инвертировать центральное кладистическое утверждение и считать кладистическую систему определяемой в конечном счете последовательностью апоморфий, а события дивергенции рассматривать как орудие определения этой последовательности. Такой подход был использован Бразерсом (Brothers, 1975) в его классическом исследовании филогении и систематики жалоносных перепончатокрылых. Сходную позицию, видимо, занимает Нельсон, утверждая, что "происхождение без изменения ... не является достаточным объяснением таксона" (Nelson 1989: 280-281).
Как дивергенция не всегда сопровождается апоморфным изменением всех ее продуктов, так и приобретение апоморфии в случае филетической эволюции оказывается не связанным с дивергенцией, а установление точной последовательности апоморфий - недостижимым даже в теории. Таким образом, два подхода по крайней мере сравнимы по их значению для филогенетического исследования - не в том смысле, что они обязательно равноценны в этом отношении, а вы том, что их достоинства и недостатки равно заслуживают специального изучения. Например, в версии Бразерса апоморфии вряд ли доступны не только точному секвенированию, но и простому подсчету: в большинстве случаев апоморфии слишком сложны, чтобы можно было представить их возникновение за один шаг (в течение одного события видообразования). Но тогда возникает вопрос об идентификации системы таксонов, иерархически организованных последовательностью элементарных шагов приобретения сложной апоморфии. С другой стороны, подход Бразерса кажется более богатым информационно, чем у Геннига, поскольку он способен учитывать не только родство, но и объем приобретенных различий (но см. ниже).
Другие, более серьезные проблемы кладизма будем обсуждать в сравнении с достоинствами и недостатками фенетики и филистики после их более подробного рассмотрения.
Итак, кладизм утверждает, что система организмов должна быть строго генеалогической, т.е. максимально точно отражать кровное родство организмов. Для этого таксоны делятся по узлам дивергенции с помощью синапоморфий, т.е. изменений, возникших у предка и выявленных у его потомков. Сходство интересно только как свидетельство генеалогии, а само по себе несущественно. Кладистический таксон не может породить другой таксон: в кладистическом мире таксонов-предков и таксонов-потомков не существует. Конечно, кроме видов, но поскольку идентифицировать предковый вид по мнению кладистов невозможно, предок - это не реальное существо или реальный вид, а просто коллекция примитивных признаков (плезиоморфий), обнаруженных у его потомков. В этом действительно есть резон, поскольку широкое распространение параллелизмов делает разговор о предковом виде в большинстве случаев беспредметным, но оно же делает иллюзорным и самое главное утверждение кладизма о единичном событии дивергенции (или, в другой его версии, о приобретении единичной апоморфии) как о единственном событии, дающем жизнь таксону.
Фенетика
В отличие от кладизма фенетика и традиционная систематика (или, в рафинированном виде, филистика) исходят из целостности живых систем и соответственно из дискретности биоразнообразия и заняты поисками гиатусов (Табл. 4). Соответственно, фундаментальная концепция должна включать процесс, генерирующий такую дискретность. Первоначально дискретность легко объяснял креационизм, сейчас на эту роль реально претендует, по-видимому, только эпигенетическая теория эволюции (см. выше).
Из трех конкурирующих подходов фенетика наиболее прямолинейна в своих целях и средствах. Поскольку дискретность биоразнобразия описывается в терминах сходства, а не родства, то по мнению фенетиков задача систематика и исчерпывается его анализом: система должна в явном виде строиться в терминах сходства, и на этом пути можно избежать субъективности с помощью исчисления сходства. Методы расчета коллективно именуются таксометрией или чаще, но менее удачно, нумерической таксономией. К сожалению, эти методы не были развиты до заявленного уровня - на мой взгляд не из-за того, что этот уровень недостижим, а потому что фенетика была вытеснена кладистикой до того, как она выполнила свою полезную работу. Действительно, методы таксометрии многочисленны и разнообразны, выбор метода, адекватного задаче, не прост и на нынешнем уровне едва ли возможен без изрядной доли субъективности. Впрочем, это проблема любого подхода, и кладизм здесь не исключение (см., например, Mickevich, 1978): выявить оптимальную область применения конкретного метода и его сильные и слабые стороны всегда можно лишь ценой изрядных усилий. Прежде чем уступить поле боя кладизму, фенетика значительно, но недостаточно продвинулась в этом направлении.
Однако коренная проблема фенетики на мой взгляд иная. Сходство, с которым она работает, это не совсем то сходство, которое требует система организмов. Фенетики работают с изученными признаками, число которых должно быть разумно велико, но не более того. Система же, чтобы оптимально выполнять свои функции, должна опираться на сходства и различия по всем признакам, изученным и не изученным, включая и те, что, возможно, не будут изучены никогда. Конечно, такое сходство недоступно прямому изучению, но оно достаточно важно, чтобы попытаться оценить его косвенно. Именно эту цель и преследует филистика.
Филистика
Филистика, как и кладизм, исходит из того, что организация живых существ определяется не только и даже не столько условиями их существования, сколько их историей. Сходство истории (кровное родство) во многом предопределяет и сходство организации. Разница в том, что методология кладизма игнорирует все, кроме родства, фенетика игнорирует родство, а филистика пытается учесть и родство, и сходство.
Филистический таксон должен быть, во-первых, фенетически максимально однороден и максимально отличен от других таксонов, во-вторых, монофилетичен. Монофилия понимается в широком смысле по сравнению с кладизмом, т.е. включая голофилию (монофилию в кладистическом смысле, см. Ashlock, 1971) и парафилию. Филогенетическая квалификация таксонов определяется прежде всего числом линий происхождения, пересекающих границы таксона: если его нижнюю границу пересекает единственная линия (один корень), таксон именуется монофилетическим (в широком смысле), если их больше - полифилетическим. Более точной, впрочем, будет другая формулировка: таксон следует считать монофилетическим, пока и поскольку нет надежных свидетельств его полифилии (презумпция монофилии таксона).
Кладисты переопределили понятии монофилии, имея в виду не только происхождение (корни) таксона, но и его дальнейшую судьбу. Если верхняя граница монофилетического (в нашем смысле) таксона не пересечена филетическими линиями, т.е. таксон не имеет явных потомков, мы его именуем голофилетическим, а кладисты - монофилетическим. Если же его верхняя граница пересечена (потомки есть), и для филистов, и для кладистов это парафилетический таксон. Кладисты именуют голофилетический таксон монофилетическим, поскольку с их точки зрения парафилетический таксон точно так же не имеет права на существование, как и полифилетический. Они не нуждаются в термине, объединяющем голофилетический и парафилетический таксоны: для них такое объединение бессмысленно. А для филистики такой термин совершенно необходим, поскольку там обе эти формы таксона (голо- и парафилетические) признаны и равноправны. Именно их объединение первоначально именовалось монофилетическим таксоном, как это и принято здесь.
Существует еще более широкое понимание монофилии- как происхождения естественного таксона несколькими корнями от общего предка в недрах предкового таксона (плюрифилия, пахифилия, монофилия в широком смысле): таковы рептилии и млекопитающие как они обсуждаются выше в связи с первой презумпцией филогенетики. Такая монофилия не только подпадает под определение полифилии в кладистической формулировке, но и вообще исчерпывает реальное содержание этого термина. Согласно определению, за его пределами остается только множественное происхождение таксона, естественность которого отрицается (мы еще будем иметь дело с ними в последней главе этой книги), но для искусственного таксона множественность его происхождения тривиальна, и вряд ли кому-то прийдет в голову специально это обсуждать. Содержать отдельный термин для этого случая мне кажется излишним, так что я буду называть полифилией все случаи множественного происхождения таксона, как это и сформулировано выше.
Таким образом, оказывается, что многие (если не все) хорошо изученные таксоны являются полифилетическими в указанном смысле. В то же время трудно усомниться в естественности рептилий, млекопитающих, членистоногих, покрытосеменных и многих других таксонов, для которых более или менее убедительно показано происхождение более чем одним корнем от предкового таксона. Таким образом, полифилия может оказаться нормой в таксономии. И тем не менее я считаю, что методологически правильнее исходить из презумпции монофилии. Тому я вижу две причины: во-первых, на нынешнем весьма низком уровне изученности большинства таксонов обнаружение множественности происхождения конкретной группы скорее будет фальсифицировать гипотезу ее естественности, чем презумпцию монофилии. Если же группа действительно естественна, то дополнительное исследование, спровоцированное обнаружением полифилии (см. ниже), только подтвердит это и в конце концов заставит признать презумпцию монофилии несправедливой в данном конкретном случае.
Вторая причина, почему отказ от презумпции монофилии (в моей формулировке, исключающей пахифилию и т. п.) мне кажется методологически опасным, состоит в том, этот отказ означает отказ от использования филогенеза для контроля естественности таксона (см. ниже) и, следовательно, переход на позиции фенетики. Мне этот переход представляется по меньшей мере преждевременным.
Вернемся к филистической процедуре. Филистический таксон был определен как монофилетический континуум (Пономаренко, Расницын, 1971). Определение монофилии дано выше, континуум же понимается как непрерывная цепь, ветвящаяся или нет, построенная из подчиненных таксонов (монофилетических континуумов меньшего объема) таким образом, что каждый из них фенетически ближе к любому из его непосредственных соседей по цепи, чем к членам любых других таксонов (Рис. 23). Таким образом, континуум отделен гиатусом (перерывом непрерывности сходства) от всех других континуумов.

Строить систему, основанную на двух разных критериях (в данном случае это родство и сходство), рискованно тем, что допускает произвол в их применении - если только сферы компетентности этих критериев четко не разведены. Кладизм и фенетика решают проблему предельно просто - отказываясь использовать тот или иной из двух критериев: кладизм игнорирует сходство, фенетика - родство. Филистика действует тоньше: она использует оба критерия, но по-разному. Сходство используется как критерий, задающий систему, родство - как критерий, контролирующий качество системы. Таксон выделяется прослеживанием гиатусов (разрывов в сходстве), а затем проверяется на единство происхождения.
Итак, филистика требует, чтобы члены ее таксона были достаточно сходны, чтобы удовлетворять критерию континуума, и при этом не обнаруживали бы признаков полифилии (происхождения от более чем одной линии предков). Если континуум оказывается полифилетичным, критерии вступают в конфликт. В этом случае систематик едва ли станет отбрасывать все, что было сделано, и начинать работу сначала. Его естественной реакцией будет попытка фальсификации этого результата путем дополнительного изучения соответствующей группы. Попытка обычно оказывается успешной в том или ином смысле. Например, более широкий анализ может обнаружить, что сходство полифилетических групп поверхностное. Отряд сетчатокрылых насекомых (Neuroptera) долгое время включал группы, близкое родство которых впоследствии не подтвердилось. Так, еще Шарп (1910) объединял под этим именем, помимо сетчатокрылообразных в современном понимании, также эмбий (Embioptera), термитов (Isoptera), сеноедов (Psocoptera), пухоедов (Mallophaga), стрекоз (Odonata), веснянок (Plecoptera), поденок (Ephemeroptera), скорпионниц (Mecoptera), и ручейников (Trichoptera). Похожей была судьба млекопитающих отряда толстокожих (Pachydermata): этот таксон долго использовался в XVIII и XIX веках для объединения слонов, тапиров, носорогов, бегемотов и свиней, но затем был отвергнут в пользу выделения более естественных (монофилетических) отрядов.
В других случаях удалось фальсифицировать гипотезу полифилии, как в случае с зайцеобразными, выделенными в свое время в особый отряд Lagomorpha, будто бы лишь конвергентно сходный с грызунами (Gidley, 1912). Хотя статус отряда за зайцеобразными сохранился, но по современным представлениям они образуют с грызунами монофилетическую группу Glires (Wilson, 1989).
Вполне возможен случай, когда обе гипотезы (сходства и полифилии) окажутся равно правдоподобными. Скорее всего это будет означать, что наша попытка была недостаточной, и нужно попытаться заново проанализировать ситуацию и устранить конфликт. Но нельзя исключить, что мы действительно добрались до реальной множественности корней естественного таксона, о которой мы уже неоднократно говорили. Как отличить одну ситуацию от другой, у меня рецепта нет, но на всякий случай лучше еще раз попытаться проанализировать систему и филогенез группы на более широком материале.
Существуют и совершенно очевидные случаи, когда традиционная систематика сохраняет несомненно полифилетические таксоны, которые не могут быть без значительных потерь заменены монофилетическими. Я имею в виду различные паратаксоны в таких областях, как палеонтология, паразитология, микология, а также многие таксоны прокариот (подробнее см. раздел о таксономической номенклатуре). Это еще раз подтверждает, что традиционная систематика в конечном счете ориентируется на сходство, а родство использует лишь как способ косвенным образом оценить тотальное сходство (в том числе по еще не изученным признакам), которое прямой оценке недоступно. Следовательно, родство здесь используется приблизительно так же, как в генниговой версии кладизма апоморфия используется для установления дивергенций, а в бразерсовой версии дивергенции служат для определения последовательности апоморфий.
Таким образом, все рассмотренные подходы одинаково последовательны (монистичны, не эклектичны): геннигов кладизм в конечном счете ориентирован на последовательность актов дивергенции, бразерсов - на последовательность апоморфий, фенетика - на наличное (доступное непосредственному изучению) сходство, филистика - на тотальное сходство (недоступное прямому наблюдению, но косвенно оцениваемое с помощью родства).
Теперь мы можем вернуться к вопросу о сравнительных достоинствах и недостатках кладизма и филистики. На мой взгляд, важнейшим достоинством филистики является ее конечная опора на сходство, т.е. на признаки как таковые. Она может удовлетвориться сходством в наличных признаках, если сколько-нибудь надежные свидетельства родства отсутствуют, и будет столь же эмпирична, как фенетика. Фактически это и будет фенетика. Но она же будет использовать все (и только!) достаточно надежные филогенетические данные, чтобы повысить надежность создаваемой ею системы, но не сделать ее слишком гипотетичной. В противоположность филистике, кладистическая система и непосредственно, и в конечном счете опирается на родство, т.е. на интерпретацию признаков (на предположения относительно эволюционной последовательности состояний признаков). В результате кладистическая система значительно более гипотетична, чем филистическая.
Другая сторона сказанного заключается в том, что филистика "всеядна", а кладизм "привередлив". Кладизм предъявляет гораздо более жесткие требования к используемым им признакам, чем филистика (и фенетика), потому что признаки, не поддающиеся надежной поляризации, бесполезны. Это касается геннигова кладизма (ручного, или "кладизма пера и бумаги"). В компьютерном кладизме можно использовать любые признаки, но если программа не сможет уверенно реконструировать последовательность изменений некоторого признака (а она вряд ли сможет это сделать много лучше, чем хороший систематик), то этот признак будет лишним в расчетах. А сверх того, признаки, хорошо работающие в какой-то части исследуемой совокупности, но обнаруживающие многочисленные параллельные изменения (гомоплазии) в других ее частях, оказываются практически бесполезными (учитываются лишь в малой степени) даже там, где в действительности они филогенетически информативны. Происходит девальвация признаков. Менее изученные таксоны (например, вымершие), охарактеризованные меньшим числом признаков, часто не находят точного места в компьютерных кладограммах, даже если сохраняют филогенетически важные признаки. В результате кладисты вынуждены объявлять паратаксонами ("плезионами") многие группы, рассматриваемые филистикой как нормальные (ортотаксоны), но известные недостаточно для того, чтобы точно охарактеризовать их родственные отношения с лучше изученными таксонами. Естественно, это снижает таксономическую эффективность кладизма.
Филистическая система, учитывающая как сходство, так и родство, более информативна, чем другие. Фаррис (Farris 1979) возражает, напоминая, что кладистическая система в ее наиболее полной форме указывает происхождение всех признаков и поэтому более информативна, чем фенетическая (и, следовательно, филистическая). Этот вывод не бесспорен, поскольку исчерпывающая полнота системы недостижима. Действительно, чем больше информации мы набираем о поляризации известных признаков, тем больше обнаруживается новых признаков, характер эволюции которых еще не известен и которые поэтому не могут быть использованы в "ручном" кладистическом анализе. В результате приходится работать с неполным набором признаков по сравнению с филистами, и в этом смысле система несет меньший объем информации. Компьютерный кладизм может использовать все доступные признаки, но поляризацию трансформационных серий он производит автоматически, исходя просто из распределения состояний признаков (по методу внешней группы). Вся прочая информация об истории признаков, широко используемая филистикой и ручным кладизмом, здесь игнорируется, так что общие потери информации велики.
Фактически любой из сравниваемых таксономических методов использует только часть доступной информации, причем эти части не слишком сильно перекрываются. Компьютерный кладизм использует обширное поле данных, но весьма избирательно и поверхностно. Фенетика действует сходным образом, но не практикует избирательности. Ручной кладизм использует ограниченный "элитный" материал, отобранный из неограниченно широкой совокупности. Филистика также "просеивает" неограниченный объем исходных данных, но она более свободна, чем "ручной" кладизм, в его сортировке, и чем фенетика и компьютерный кладизм, в его интерпретации. Поэтому она способна к извлечению более обширной и разнообразной информации, чем другие подходы.
Существует и другой аспект этой проблемы. Из сказанного выше видно, что информация заключена не в системе самой по себе, а в ее описании или, скорее, в том, что лежит в ее основании. Система как таковая содержит только весьма ограниченную информацию о топологии соответствующей дендрограммы и о длине ее междоузлий, интерпретированной в таксономических рангах. Так, симметричная дихотомическая дендрограмма о четырех конечных таксонах и без дополнительной информации о их рангах содержит три бита информации и не больше. Детализация ранжирования таксонов обогащает информационное содержание системы, и в этом плане наиболее богаты оригинальные системы Геннига с их невероятным числом нумерованных рангов, исчерпывающе отражающих предполагаемую последовательность актов дивергенции. Однако этот подход был признан непрактичным, и он практически оставлен в пользу традиционной линнеевой иерархии рангов (Wiley, 1979). Такая форма ранжирования содержит ограниченную кладистическую информацию и использует ее произвольным образом (привязка ранга к одной из многих последовательных дивергенций произвольна), делая тем самым послегенниговый кладизм эклектичным. Искусственные правила типа нельсонового phyletic sequencing (Nelson, 1973b; Cracraft, 1974) мало помогают. Мы можем договориться, что гребенчатую кладограмму следует преобразовать в группу таксонов одного ранга, перечисляемых в порядке их дивергенции, начиная с отошедшего первым, но в последней паре порядок неизбежно будет произвольным. И мы не сможем отличить такой список от другого, построенного произвольно или основанного на более сложной кладограмме, не строго гребенчатой или/и включающей политомию.
Еще более резкое отклонение от базового кладистического принципа представляет популярное различение crown group (эквивалент *таксона Геннига) и stem group (эквивалент таксона минус *таксон в схеме Геннига). Действительно, stem group это по определению парафилетический таксон, запрещенный в кладизме.
Утверждается, что фенетика и филистика в отличие от кладизма используют таксоны, охарактеризованные отсутствием признака (Platnick, 1979), именно парафилетические таксоны, характеризующиеся только симплезиоморфиями, т.е. отсутствием апоморфий. Выше, однако, было показано, что апоморфия не есть признак или состояние признака. Апоморфия - это гипотеза относительно истории признака, так что отсутствие апоморфии не означает отсутствия признака. Как уже упоминалось, ящерицы в отличие от змей действительно лишены апоморфии, зато у них есть признак - ноги.
Заслуживает обсуждения также проблема гибридных и симбиотических таксонов, важная в силу хотя бы того, что они слишком обычны, чтобы их игнорировать как нечто экзотическое. Первую категорию представляет не менее чем сам таксон эвкариот, а гибриды очень характерны, например, для многих таксонов растений. Согласно обзору Цвелева (1993), в семействе злаков (Poaceae) триба Triticeae включает 500 видов, из которых 300 кариотипически определенно представляют собой межродовые гибриды, которыми, в частности, составлены самые большие роды. Утверждается, что то же свойственно и многим другим таксонам высших растений. Симбиотический и гибридный таксоны полифилетичны (или составляют часть полифилетичного таксона) по определению, поскольку их граница (или граница старшего таксона) пересечена более чем одной предковой линией, что категорически нарушают принципы кладизма. Тем не менее такие таксоны принципиально не могут быть расчленены на голофилетические составляющие и поэтому должны быть исключены из компетенции кладизма, с чем вряд ли кто-либо согласится.
Существуют и другие соображения, имеющие отношение к сравнению конкурирующих таксономических концепций. Однако и сказанного достаточно для заключения, что каждому из них свойственны свои достоинства и недостатки, которые, впрочем, не всегда уравновешивают друг друга.
Кладизм сводит систему группы к ее генеалогической истории, фенетика к доступным для анализа признакам сходства, а традиционная систематика (в зеркале филистики) пытается использовать и то, и другое, причем не произвольно, а упорядоченным образом. При этом, если сколько-нибудь надежные свидетельства родства отсутствуют, традиционная систематика удовлетворится просто сходством в наличных признаках, и тогда это будет фенетика. Но когда возможно, она же будет использовать все доступные филогенетические данные, чтобы повысить надежность создаваемой ею системы.
Фенетика сугубо эмпирична, а кладизм, напротив, высоко гипотетичен, так как и непосредственно, и в конечном счете опирается на родство, т.е. на интерпретацию сходства. Филистика же свободна занимать любую позицию в этом спектре в зависимости от свойств материала, доступного в каждом отдельном случае. Филистика "всеядна" и может работать с любым доступным материалом, используя при необходимости любые фенетические и кладистические методы. Напротив, кладизм "привередлив" и предъявляет гораздо более жесткие требования к используемым им признакам. Поэтому так часто кладистическая система оказывается либо неустойчивой, когда минимальные изменения параметров расчета существенно меняют форму кладограммы, либо неинтересной ("не разрешенной", с не выясненными отношениями многих таксонов) (Рис. 19). По этой же причине кладисты вынуждены объявлять паратаксонами ("плезионами") многие группы, которые для филистики вполне нормальны.
Тем не менее кладизм - это доминирующая система взглядов на Западе, и у нас дело, кажется, идет к тому же. Так проще. Четкая методика, меньше места для интуитивных оценочных функций, вроде оценки уровня сходств и различий, нет отдельных задач построения системы и филогенеза. Особенно велики эти преимущества у компьютерного (экономного) кладизма. Здесь сохраняется единственный этап, на котором еще нужно думать: это выделение группы для анализа, поиск и отбор признаков для расчета кладограммы. Впрочем, кладисты не утруждают себя анализом методологии этой важнейшей части работы. А дальше просто: составляем матрицу, вставляем ее в сложную, но детально описанную и потому вполне доступную для пользователя компьютерную программу, запускаем мощный компьютер и ждем окончания расчетов. Иногда приходится ждать долго: пять лет назад компьютер в Швеции обсчитывал мои данные больше месяца. Если кладограмма не понравилась, можно изменить параметры расчета, и пусть компьютер посчитает еще. В конце концов он выдаст приемлемый результат, хотя бы минимально согласующийся со здравым смыслом (т.е. с априорными ожиданиями).
В общем, на мой взгляд, все-таки лучше заниматься традиционной систематикой, но при этом четко понимать ее основания, критерии и цели, сформулированные филистикой, и использовать методы кладизма и фенетики везде, где они могут помочь.
ТАКСОНОМИЧЕСКАЯ НОМЕНКЛАТУРА
Номенклатура - это совокупность договоренностей о том, как создавать и использовать названия таксонов. Обычно она воспринимается как область усложненных деталей и простых до очевидности базовых принципов. Это неверно, во-первых, поскольку еще далеко не все действующие принципы биологической номенклатуры осознаны и сформулированы в явном виде. Некоторые из них упомянуты в кодексах (см. МКБН, 1985; МКЗН, 1988, 2000), но не в качестве базовых принципов, других там просто нет. Во-вторых, некоторые из базовых принципов не являются чистыми соглашениями, а отражают определенные таксономические концепции или представляют следствия из определенных эволюционных гипотез. Линнеев принцип биноминальной номенклатуры вполне конвенционален и потому легко сочетается с любой таксономической концепцией, признающей иерархию родов и видов. Иное дело принцип типа, как это подробно обсуждается далее. Различение этих двух типов принципов представляется полезным, и в дальнейшем они будут именоваться таксономически независимыми и таксономически зависимыми принципами соответственно (Табл. 5).
Таблица 5. Принципы номенклатуры
|
таксономически независимые |
||
|
1 |
невмешательства в принятие таксономических решений за пределами ограничений, налагаемых таксономически зависимыми принципами |
|
|
2 |
ограниченной сферы приложения |
|
|
3 |
гомонимии |
|
|
4 |
приоритета |
|
|
5 |
стандартизированных, ранг-специфичных названий |
|
|
6 |
верховного приоритета Международной Комиссии по номенклатуре |
|
|
таксономически зависимые |
||
|
7 |
типа (нет разумных причин отвергать) |
|
|
8 |
иерархии (соблюдается всегда) |
|
|
9 |
синонимии (применение к паратаксонам ограничено рамками соответствующей частной системы) |
|
Мой личный опыт заставляет меня анализировать преимущественно зоологическую номенклатуру, но это не столь существенно, поскольку соответствующие кодексы различаются скорее деталями, чем базовыми установками.
Таксономически независимые принципы
Таксономически независимые принципы обычно просты, не требуют больших комментариев и здесь рассматриваются первыми.
1. В действующем Кодексе сказано, что он "воздерживается от вмешательства в таксономические суждения, которые не могут подвергаться регулированию" (МКЗН 2000, с. 24). Это утверждение не вполне справедливо из-за существования таксономически зависимых принципов, которые не позволяют принимать некоторые таксономические решения. Поэтому формулировка должна быть дополнена словами "за пределами ограничений, налагаемых Кодексом ".
2. В зоологии область применения принципов номенклатуры ограничена надпопуляционными группами от подвида до надсемейства (подробнее см. МКЗН 2000, ст. 1.2). Исключение старших таксонов из сферы действия кодекса на мой взгляд ошибочно, но это предмет особой дискуссии (Rasnitsyn, 1982, 1989, 1991, 2002b). Вкратце эти соображения можно изложить следующим образом.
Обеспечиваемая кодексами стандартизация образования и использования названий доказала долгой практикой свое благотворное влияние на всю биологию. Отказ от ее распространения на старшие таксоны вряд ли связан с их глубинной спецификой: серьезных указаний на такую специфику не известно. Главное достижение биологической номенклатуры - принцип типа, соответствующий природе таксона (см. ниже). Этим обеспечивается устойчивость употребления и упорядоченность изменения названия при самых различных изменениях состава (границ) и интерпретации таксона (ранг, признаки и т. п.). Дело, конечно, именно в типификации, так как попытки стандартизировать нетипифицированные названия старших таксонов столь многочисленны, что снимают все сомнения в том, что потребность в такой унификации не только существует, но и широко осознана.
Первые попытки типификации старших таксонов насчитывают уже более двух веков (Laicharting, 1781), и понимание ее необходимости также растет. Эта точка зрения победила в ботанике (МКБН, 1985) и находит некоторую поддержку в зоологии (Родендорф, 1977, Starobogatov, 1991, Клюге, 2000). Дело в том, что нетипифицированное название отнесено к таксону как к целому и практично только до тех пор, пока интерпретация самого таксона остается устойчивой. В противном случае возникают проблемы с применением названия, что выливается в разнобой и переименования. Однако введение типифицированных названий старших таксонов и, соответственно, их переименование затрагивает интересы многочисленных пользователей, которых мало интересуют проблемы систематики высоких уровней, но пугает перспектива заучивать новые слова. Для тех же, кто работает со старшими таксонами, это беда: таксономические и филогенетические ошибки и неточности консервируются нетипифицированными названиями и накапливаются. Чем дольше это продолжается, тем труднее будет переходить к типификации старших таксонов. Но переходить все равно придется, и потомки не скажут нам спасибо за наш эгоистичный консерватизм.
3. Идентичные названия разных таксонов не должны использоваться в качестве действительных названий (МКЗН 2000, ст. 52). Область применения принципа гомонимии произвольно ограничена таким образом, чтобы не затрагивать случаев гомонимии, во-первых, между названиями видов в разных родах, во-вторых, между таксонами растений и животных, и в-третьих, с и между таксонами рангом выше надсемейства.
4. Выбор между конкурирующими названиями (синонимами или гомонимами) должен осуществляться в пользу того названия, которое было предложено раньше (МКЗН 2000, ст. 23). Это принцип приоритета.
5. Принцип стандартизированной и ранг-специфичной формы названий таксонов. Этот принцип представляет основу для многочисленных правил для языка, грамматики и синтаксиса названий, и включает принцип биноминальной номенклатуры (МКЗН 2000, ст. 4-6, 11, 25-34).
6. Последний среди таксономически независимых принципов - принцип верховенства Международной Комиссии по номенклатуре, которая может принимать решения в нарушение любых положений Кодекса, кроме тех, которые касаются ее самой (МКЗН 2000: ст. 78).
Таксономически зависимые принципы
Таксономически зависимых принципов три:
1. Принцип типа. Это самый важный из таксономически зависимых принципов, и на его примере хорошо видны те ограничения, которые таксономическая концепция может накладывать на номенклатуру. В Кодексе 1988 г. этот принцип сформулирован следующим образом: "Номенклатурный тип служит объективным эталоном, с помощью которого приложение названия определяется независимо от возможных изменений границ таксона" (МКЗН 1988, ст. 61а; формулировка МКЗН 2000, ст. 61.1, кажется менее удачной: "Фиксация номенклатурного типа номинального таксона обеспечивает объективный эталон для приложения названия этого таксона"). Другими словами, таксон не может быть введен в систему иначе как, в конечном счете, ссылкой на тип, несущий его название. Чтобы оценить смысл этого утверждения, рассмотрим альтернативные возможности.
Первое, что приходит в голову, это возможность введения таксона в систему ссылкой на его признаки. Это означает, что признаки таксона выполняют определяющую, а не просто диагностирующую функцию, а сам таксон соответствует понятию "класс" (Ghiselin, 1974, 1987). Существуют разные возможности такого введения таксона в систему, простейший из которых - комбинаторная система, т.е. многомерная матрица, каждая ячейка которой соответствует определенной комбинации признаков и является местом таксона, обладающего такой комбинацией. Версию такой матрицы представляет политомическая таблица, иногда используемая систематиками, т.е. прямоугольная матрица с рядами, представляющими таксоны, и колонками, соответствующими признакам (как на табл. 6). В результате каждая ячейка отражает состояние одного конкретного признака в определенном таксоне. Эта форма системы действительно проста и иногда очень полезна, но не соответствует задачам общей системы организмов. Она не обладает гибкостью: мы не можем исправить ее локально, чтобы, например, лучше согласовать ее с признаками конкретного таксона, поскольку введение нового, переопределение или удаление старого признака заставляет пересматривать характеристики всех таксонов. Поэтому даже в качестве средства идентификации комбинативная система используется нечасто.
Таблица 6

Более гибка система, в которой таксоны определяются ранжированными признаками, как обычная определительная таблица, в которой чем выше ранг, который мы приписываем признаку, тем раньше от появляется в таблице. Эта система действительно удобна, хотя и не столь хороша, как общая система организмов, из-за очевидно произвольного ранжирования признаков. Тем не менее в несколько модифицированной форме (с выделением небольшого числа высокоранговых признаков, достаточного для формирования системы) этот подход был весьма популярен у систематиков, начиная с Линнея (Linnaeus, 1751) с его упором на признаки фруктификаций у растений. В наиболее четкой форме этот подход был сформулирован Любищевым (1923, 1966), призывавшим к поиску немногих наиболее высокоранговых признаков (параметров), детерминирующих распределение всех других признаков, - подобно ядерному заряду, который определяет свойства атомов и, соответственно, положение элементов в таблице Менделеева. Результатом должно было стать открытие параметрической системы организмов, позволяющей прогнозировать все важные свойства соответствующих таксонов по этим ключевым признакам (параметрам). Поставленная Любищевым задача казалась безнадежной, и самому Любищеву не удалось ее разрешить. Тем не менее решение в каком-то смысле было найдено, но совсем не там, где он его искал. По иронии судьбы это была область, которую сам Любищев как убежденный антиселекционист считал ложной.
Эта область - кладистическая система с ее центральным утверждением, что признаки организма и, соответственно, его положение в системе наилучшим образом определяются единственным признаком - родством. Родство оказалось любищевским параметром по определению. Смысл и возможности кладистической системы обсуждены выше, здесь я хочу только привлечь внимание к тому факту, что эта система не нуждается в типификации ее таксонов, поскольку ссылка на значение параметра (характер родственных отношений) достаточна для введения таксона в систему.
Действительно, "названия синонимичны, если они указывают на клады [ветви], отходящие от одного и того же предка" de Queiroz & Gauthier, 1990: 307). Авторы этого утверждения не сделали из него самоочевидный вывод, что кладизму следует отказаться от принципа типа, но этот шаг сделали Сандберг и Плейел (Sundberg & Pleijel 1994). Мне не известно о практическом применении их предложения, и я боюсь, что это будет трудная задача. Предложение состоит в том, что название таксона должно быть подколото подобно этикетке не к типу, а к предку соответствующей клады. Однако кладистический подход, как мы могли видеть, не позволяет рассматривать предка в качестве реальной группы, которую можно идентифицировать и изучать. Напротив, предок для кладиста это только сумма синапоморфий данной клады плюс то же для всех клад (таксонов), включающих данную. Все эти синапоморфии представляют собой не результат наблюдения, как типовой экземпляр, а гипотезы о состояниях признака, приобретенных в соответствующих актах дивергенции. Таким образом, нам предлагают подкалывать имя таксона вместо коллекционного экземпляра к коллекции гипотез. Я не думаю, что систематиков будут легко убедить принять этот метод типификации.
Логической альтернативой класса как группы, определяемой ее признаками, является индивидуум, и предложение считать таксон индивидуумом (Ghiselin, 1974, 1987 и приведенная там библиография) вполне естественно. Обладая свойствами индивидуума, таксон может вводится в систему остенсивно, т.е. прямым указанием (в том числе и ссылкой на его название). Для этого его целостность в пространстве и во времени должна быть достаточно высокой, чтобы позволять таксону рождаться и умирать, но не распадаться на части, которые могли бы претендовать на наследование имени таксона. Пусть человек на пути от младенца к старику не сохранил ни одного общего атома вещества и ни одного общего признака, но при всех его метаморфозах всегда можно проследить, что это одно лицо. Индивидуум настолько целостен, что использование его имени не вызывает проблем. Можно просто указать пальцем - вот это, и не детализировать, к чему именно относится имя - к голове, сердцу, уху или чему-то еще. Ко всему сразу и к каждой детали в отдельности. Именно здесь проходит граница между остенсивным методом и принципом типа: если любая часть индивидуума равно является носителем его названия, специальные правила выбора и использования номенклатурного типа, служащего для введения таксона в систему, оказываются излишними.
Если метод типа не приложим к индивидууму, то таксон - это не индивидуум. По крайней мере он не типичный индивидуум (см., например, комментарии к статье Ghiselin, 1981), так что проблема требует дальнейшего обсуждения. Парадигмальный пример индивидуума, организм, целостен из-за непрерывного взаимодействия его частей. То же самое утверждалось в отношении вида, целостность которого обеспечена обменом генами (майрова концепция биологического вида). Однако эта модель имеет ограниченное приложение, хотя бы потому, что "Концепция биологического вида применима лишь к тому, что я назвал 'непротяженной ситуацией', когда популяции вида находятся в реальном [репродуктивном] контакте" (Mayr, 1988: 301-302). На самом деле проблем здесь, как мы уже видели, еще больше (см. раздел об онтологии эволюции).
Однако дихотомия класс - индивидуум не исчерпывает всех возможностей. Таксон обладает чертами обоих этих крайностей. В качестве класса он обладает и признаками, о чем свидетельствует наличие диагноза, и членами (вопреки Гислину, я являюсь не только частью вида Homo sapiens, но и его экземпляром, т.е. примером, представителем). В качестве индивидуума таксон обладает частями (популяции по отношению к виду) и, что важнее, способностью развиваться (эволюционировать), не теряя своей индивидуальности, т.е. сохраняя свою целостность во времени, в многомерном пространстве признаков и, в той или иной степени, в географическом пространстве. Именно эта целостность позволяет идентифицировать таксон как индивидуум, т.е. по его имени. Обладая признаками класса и индивидуума, таксон занимает промежуточное положение, а точнее, заполняет пространство между ними, превращая класс и индивидуум в два полюса единого спектра.
Таким образом, таксон - это нечто промежуточное между классом и индивидуумом. Он достаточно целостен, чтобы, меняясь, сохранять имя как средство опознания, но недостаточно целостен, чтобы простого "вот оно" было достаточно для его опознания. Недостаточно целостен, потому что таксон - это континуум, сгущение точек, облако в многомерном пространстве признаков (Рис. 24). Облако, которое может эволюционировать, т.е. менять свои размеры и положение в пространстве признаков, может исчезнуть (вымереть) или, наоборот, разделится пополам либо отпочковать новое облако, и т. д. Континуум характеризуется как его целостностью, выявляемой по наличию гиатуса (нарушения непрерывности) между ним и другими облаками-континуумами, так и признаками его диагноза. Однако оба эти критерия не обнаруживают жесткости и легко изменяются как в ходе эволюции, так и по мере накопления наших знаний о таксоне, и в любой момент может обнаружиться их неспособность четко идентифицировать и разграничить таксоны.

Возможность идентификации континуума, в отличие от индивидуума, определяется сходством, но в отличие от класса речь идет об отношении (общем сходстве), а не о сходстве по конкретным признакам. Поэтому указание пальцем не удается адресовать ни облаку в целом, ни его конкретным признакам. Оно должно быть адресовано определенной точке этого облака - его номенклатурному типу. Что бы ни произошло с таксоном-облаком - в ходе ли его эволюции, или из-за меняющихся наших представлениях о нем, но имя, закрепленное за номенклатурным типом, как этикетка за музейным экземпляром, всегда покажет нам, о чем идет речь. Если, конечно, тип в сохранности - или хотя бы описан с приемлемой полнотой.
Сказанное делает понятным, что номенклатурный тип - это естественный атрибут традиционной, т.е. филистической, а также фенетической системы, и, соответственно, выводится из эпигенетической теории эволюции. Что же касается кладизма, то кладистический таксон определяется, как мы видели, ссылкой на событие дивергенции или на синапоморфию, приобретенную этим таксоном (его первым членом) в момент возникновения. Именно они (строго говоря, одно из них) мыслятся как событие, создающее таксон. Это событие есть признак, единолично и однозначно определяющий кладистический таксон. Таким образом, кладистический таксон - это класс по определению, и для его идентификации не нужен никакой номенклатурный тип: все, что необходимо и достаточно, это, как уже отмечалось, акт дивергенции и/или синапоморфия.
К тому же результату можно прийти, используя иной и более короткий путь рассуждений. Цель любой классификации состоит в том, чтобы покрыть таксонами все подлежащее классифицированию разнообразие без остатка и, по возможности, без перекрывания границ таксонов. Если мы принимаем, что таксоны как естественные группировки и образуемая ими система существует в природе, а не создана систематиками (есть и такая точка зрения, см., например, С. Расницын, 2000), главную цель классификации составляет, во-первых, прослеживание естественных перерывов непрерывности в классифицируемом пространстве (пользуясь платоновой метафорой, цитированной Hull, 1983: 186 - расчленять природу по ее сочленениям), во-вторых, фиксация и ранжирование выявленных гиатусов в качестве таксономических границ (при искусственной классификации мы налагаем эти границы вместо того, чтобы прослеживать их). Классы не поддаются такой процедуре, так как они определяются признаками, для которых естественно перекрываться с другими признаками (если только не определять класс так изощренно, как это я сделал выше, анализируя природу кладистического таксона). Что касается индивидуумов, нам не известно природных сил и закономерностей, которые могли бы упаковать индивидуумы плотно, не оставляя свободных промежутков (опять-таки, если не видеть в кладистическом таксоне индивидуум на основании того, что он рождается с дивергенцией своего предка и впоследствии может вымереть). Таксон-континуум, напротив, идеально соответствует указанной процедуре классификации, поскольку единственный законный способ определения континуума - прослеживание и ранжирование его границ.
Помимо принципа типа, существуют еще два таксономически зависимых принципа номенклатуры, которые, как и принцип типа, следуют из континуальной природы таксона.
8. Принцип иерархии. Будучи континуумами, таксоны не могут быть объединены иначе как в следующие более широкие континуумы (старшие таксоны), которые должны покрывать включаемые континуумы целиком. Равным образом таксон-континуум не может одновременно входить в два и более старших таксона не целиком, ни частями, так как иначе эти старшие таксоны будут перекрываться, нарушая тем самым определение континуума. Все это означает, что система имеет строго иерархическую форму.
9. Принцип синонимии. Если таксон может быть членом единственного старшего таксона, он имеет единственное место в системе и, следовательно, ему достаточно одного имени. Все другие названия должны быть признаны недействительными, т.е. быть объявлены синонимами.
Таксоны, нарушающие принципы номенклатуры
Жизнь сложна, и наука тоже. Иногда нам приходится нарушать принципы номенклатуры - не по невежеству или разгильдяйству, а по необходимости, когда для этого возникают серьезные причины. Рассмотрим эти возможные причины.
Таксономически независимые принципы представляют собой соглашения между систематиками и в этом смысле субъективны. Причины отказа следовать им тоже всегда субъективны в том смысле, что они не оправданы какими-то таксономическими соображениями. Поэтому они здесь не рассматриваются. Проблема таксономически зависимых принципов более важна и заслуживает анализа. Более детально такой анализ проведен ранее (Расницын, 1986а) и здесь дается в сокращенном виде.
Нарушать таксономически зависимые принципы номенклатуры нам приходится, когда недостаток информации, необходимой для классификации некоего таксономического материала, вступает в конфликт с необходимостью его тем не менее классифицировать - из-за научной ли или прикладной его значимости. Неполнота информации в этих случаях обычно связана со специфической дефектностью материала, как это хорошо известно в палеонтологии и в систематике групп с усложненным онтогенезом (например, у паразитических червей и грибов).
Международные кодексы номенклатуры признают некоторые формы таксонов особого статуса, но не решают проблему в целом, а содержащиеся в кодексах регламентации во многом противоречивы. Неудовлетворительность ситуации с таксонами особого статуса трудно считать неожиданной по крайней мере в зоологии, где такие таксоны традиционно считаются чем-то редким и крайне нежелательным. В целом (как в зоологии, так и в ботанике) они действительно не слишком многочисленны, но в некоторых специфических областях (палеонтология, паразитология, микология) отказ от их использования грозит большой потерей информации об объектах, имеющих важное стратиграфическое, медицинское или агро-, лесохозяйственное значение. Использование "неполноценных" таксонов неизбежно, но при неполной регламентации кодексами это ведет к номенклатурному разнобою и самодеятельности. В палеонтологии, например, в принципе однотипные группировки в одних случаях обозначают как специальные таксономические категории (ихнотаксоны в ихнологии, в частности "индузироды" и "индузиподроды" в систематике ископаемых домиков, построенных личинками насекомых - ручейников (Вялов, Сукачева, 1976), турмы, антетурмы и т.п. в палеопалинологии, формальные роды и недавно отмененные орган-роды в систематике макроостатков растений, и т. д.). В других случаях берут в кавычки название таксона (Белокрыс, 1984) либо таксономической категории (Пономаренко, 1985), в третьих - просто оговаривают статус таксона в описании или преамбуле. Порой дело доходит до разработки специальных кодексов номенклатуры (Sarjeant, Kennedy, 1973).
Этот разнобой неудобен, хотя и не опасен. Более того, он даже полезен как способ поиска оптимальных форм работы с "неполноценными" таксонами. Опасно порожденное негативным отношением к таким таксонам желание представить их нормальными, полноценными. Удивительно, например, стремление многих специалистов по конодонтам (изолированным микроскопическим элементам скелета древних животных), показать полноценными их таксоны (Melville, 1981a,b). Еще более демонстративна ситуация с современным родом Laberius Kieffer (перепончатокрылые насекомые семейства Dryinidae). Этот род был описан (под названием Labeo Hal., nom. praeocc.) как обычный таксон, но затем был использован как сборная группа для самцов триб Dryinini и Gonatopodini (Kiefer, 1914). Эти трибы характеризуются сильным половым диморфизмом, причем их система построена по признакам самок, а самцов, если они не ассоциированы с самками экспериментально, до последнего времени не удавалось определить даже до трибы. Однако в последней ревизии семейства (Olmi, 1984) ее автор отказался от такой интерпретации рода Laberius и свел его в синонимы рода Dicondylus Haliday (не обсуждая при этом признаки типового вида Laberius, т. е. не обосновывая своего решения). При этом вымерший вид, описанный в составе рода Laberius, понимаемого как сборная группа (Н. Пономаренко, 1981), был переведен в род Dryinus Latreille с указанием, что единственный экземпляр этого вида сохранился "настолько плохо, что это не позволяет отнести его к какому-либо роду семейства Dryinidae" (Olmi, 1984). Тем самым Dryinus (типовой род семейства) был превращен фактически в сборную группу, т.е. самый неполноценный таксон (см. ниже), хотя цель автора была явно обратной - избавиться от неполноценного таксона.
Таким образом, проблема таксонов, не удовлетворяющих тем или иным правилам номенклатуры, остается актуальной. Проанализируем более детально природу и формы таких таксонов (Табл. 7).
Таблица 7. Формы таксонов
|
таксон |
принадлежит системе |
тип и диагноз |
при синонимии |
|
ортотаксон |
общей |
собственный |
подлежит синонимизации |
|
неясного положения |
общей |
собственный |
подлежит синонимизации |
|
формальный (паратаксон) |
специальной |
собственный |
не подлежит синонимизации |
|
сборный (паратаксон) |
специальной |
старшего таксона |
не подлежит синонимизации |
Самый легкий случай - это таксон неясного положения (taxon incertae sedis). Он отличается от нормального таксона (ортотаксона) тем, что неполнота его характеристики не позволяет уточнить его положение только на определенном уровне иерархии. Например, род неясного положения (genus incertae sedis) на данный момент может быть с достаточной уверенностью отнесен к определенному отряду, но не семейству. Вымерший род Baissobius Rasn., описанный среди перепончатокрылых насекомых подотряда Apocrita, "обладает своеобразным типом редуцированного жилкования, не встречающимся, насколько известно, у других Hymenoptera" (Расницын, 1975: 128), причем его жилкование достаточно своеобразно, чтобы затруднить определение систематического положения насекомого (другие важные признаки на ископаемом не видны), но оно недостаточно своеобразно, чтобы оправдать выделение особого семейства (впоследствии дополнительные находки позволили расстаться с этим таксоном неясного положения, превратив его в нормальный род, принадлежащий семейству Embolemidae; Rasnitsyn, 1996b).
Использование таксонов неясного положения можно понять как локальный отказ от принципа синонимии. Действительно, род, подчиненный отряду, минуя семейство, фактически выступает в качестве самостоятельного, но не названного семейства, скорее всего синонимичного какому-то из существующих семейств, только пока неизвестно, какому именно. Как видно из последнего примера, со временем эта синонимия действительно может стать известной.
Более сложные случаи, объединенные понятием паратаксономии, затрагивают принципы синонимии и иерархии. Определение, трактующее паратаксон как таксон в частных системах для изолированных органов и частей тела животных (Melville, 1979), уже практического применения этого понятия. Действительно, специфика и проблемы, связанные с введением и использованием таксономического названия, мало отличаются в частных системах, созданных для изолированных органов и частей тела организмов, для определенного пола или стадии жизненного цикла, или для следов жизнедеятельности. Поэтому здесь принято другое определение: паратаксон в отличие от ортотаксона и таксона неясного положения принадлежит самостоятельной системе, связанной с основной системой принципом гомонимии, но не принципом синонимии, и потому не подлежит синонимизации даже в тех случаях, когда показана его синонимия с тем или иным ортотаксоном. Таким образом, паратаксон - это таксон, на который не распространяется требование уникальности таксономического положения, т.е. требование вхождения в единую для всех организмов иерархию. Именно к этому, в частности, сводятся все пять признаков своеобразия ископаемого материала, сформулированные С.В. Мейеном (Meyen, Traverse, 1979; Мейен, 1983). Важные соображения о природе и функциях паратаксона приведены Мейеном (1990).
Различают несколько видов паратаксонов. Термин формальный таксон было предложено (Расницын 1986а) использовать как более широкую интерпретацию ботанического понятия форм-рода. Формальный таксон - это эквивалент нормального таксона (ортотаксона), но используемый только в рамках специальной системы. Эта система параллельна как основной системе, так и другим таким же частным системам, но независима от них в отношении принципа синонимии. Таковы системы изолированных ископаемых листьев, семян или жучьих надкрыльев, ископаемых следов передвижения организмов и других форм их жизнедеятельности (например, домиков личинок ручейников), система личиночных стадий современных паразитических червей или не ассоциированных с самками самцов в некоторых группах насекомых, где система ориентирована на признаки самок. Формальный таксон отличается от ортотаксона только тем, что может быть, а порой точно является синонимом какого-то ортотаксона, и там не менее он не подлежит синонимизации. Хорошо известно, например, что в ряде случаев для разрозненных частей древних растений (листьев, стволов, корней, мужских и женских фруктификаций, семян и пыльцы), описанных каждая в своей специальной системе, известны также находки в прижизненной связи, позволяющие "собрать" целое растение (рис. 25). Тем не менее каждая из таких частей сохраняет свое название, полученное в качестве формального таксона. Так, хорошо сохранившиеся карбоновые стволы Lepidodendron, стволы Aspidiaria, Bergeria, Aspidiopsis, Knorria с корой, опавшей на разную глубину, корни Stigmaria, женские стробилы Lepidocarpon, мужские стробилы Lepidostrobus и микроспоры Lycospora (рис. 26) могли принадлежать одному и тому же растению, поскольку прижизненное сочетание разных пар из этого списка более или менее надежно установлено (Мейен, 1987б). И никто при этом не покушается на валидность всех этих родовых названий!
Относительно некоторых случаев этого рода утверждается, что они затрагивают и принцип типа. Иногда резоны таких утверждений трудно понять (как в случае отказа типифицировать ихнотаксоны, т.е. таксоны системы ископаемых следов, в версии МКЗН 1988, ст. 66), в других причины подробно обсуждаются. А.Г. Пономаренко (1985) установил нетипифицированные формальные таксоны, определяемые только признаками их диагноза ("Род [Flichea Handlirsch.- А.Р.] понимается как формальное объединение изолированных надкрылий жуков без отчетливых борозд на верхней стороне и с заметной вырезкой посредине наружного края", с. 76]). Как объяснил мне автор, причиной явилось упорное стремление некоторых систематиков синонимизировать и таким образом ликвидировать паратаксоны, сколь произвольной ни была бы такая синонимизация, и в частности упомянутый выше пример с родом Laberius Kieffer. Нетипифицированные таксоны Пономаренко - это, по существу, те же формальные таксоны, но искусственно детипифицированные и тем самым выведенные из-под юрисдикции кодекса номенклатуры (поскольку кодекс регламентирует применение названия ссылкой на тип, см. выше). Детипификация действительно предотвращает необоснованную синонимизацию, но проблема в том, что она исключает всякую синонимизацию и поэтому оказывается лекарством хуже, чем сама болезнь. Мне кажется, что понимание целей и функций паратаксономии будет лучшей защитой от произвольной синонимизации.

Рис. 25. Взаимное соответствие родовых названий для (сверху и по часовой стрелке) пыльцы, семян, женских фруктификаций, листьев и мужских фруктификаций семенных папоротников семейства Peltaspermaceae. Соответствие основано на находке отдельных представителей соответствующих родов в их прижизненной связи, т.е. когда разные органы найденного ископаемого растения по своим признакам оказываются соответствующими тем или иным из родов, показанных на схеме (по Meyen, 1987).

Рис. 26. Формальные таксоны, предложенные для классификации изолированных частей древних лепидофитов (по Мейену, 1987б, 1988): Lepidodendron для стволов с сохранившейся корой, Aspidiaria, Bergeria, Aspidiopsis, Knorria для стволов, в разной степени утративших кору, Stigmaria для корней, Lepidocarpon для женских стробилов (показан общий вид и разрез мегаспорангия), и микроспора Lycospora, извлеченная из микроспорангия Lepidostrobus. Все эти названия в некоторых случаях обозначают одно и то же растение, части которого в разных сочетаниях неоднократно были найдены в прижизненной связи (как на фото слева внизу).
Последний и самый бедный содержанием вид паратаксона это сборный таксон (collective taxon). Это паратаксон, обычно в ранге рода, который отнесен к определенному старшему таксону, но в отличие от формального таксона он по бедности своей характеристики не может быть организован там в специальную систему, параллельную основной. По существу, виды в сборных группах - это просто виды неясного положения (species incertae sedis). Например, название "Cercaria O.F. Müller, 1773 установлено для рода червей, и многие авторы в XIX в. обращались с ним как с названием номинального рода, как если бы его типовым видом был C. lemna O.F. Müller, 1773. В настоящее время Cercaria употребляется как название сборной группы для личинок трематод, которые не могут быть с уверенностью отнесены к нормальным родам" (МКЗН 2000, ст. 67.14). Различие между таксами неясного положения и видами сборного рода состоит только в том, что вид не вправе появляться в одиночку, без названия рода. Поэтому для таких видов приходится вводить название сборной группы, которое имеет вид родового названия, но выполняет роль названия того старшего таксона, к которому автор относит виды, включенные в данный сборный род. Из ранее упоминавшихся случаев сюда относятся, кроме Cercaria, также Laberius в интерпретации Кифера (Kieffer, 1914), т.е. эквивалент Dryinini + Gonatopodini, и Dryinus в интерпретации Ольми (Olmi, 1984), т.е. эквивалент Dryinidae.
Сборные таксоны считаются детипифицированными, но это утверждение ошибочно. Приведенное выше определение означает, что сборный таксон по характеристике и объему совпадает с соответствующим старшим таксоном, и его члены, подобно ортотаксонам, вводятся в систему в конечном счете ссылкой на номенклатурный тип последнего. Виды Cercaria определимы как таковые просто в силу того, что достаточно сходны с личинками типового вида трематод. А то, что типовой вид трематод пока формально не зафиксирован, несущественно: я не сомневаюсь, что старшие таксоны будут типифицированы и в зоологии. Другой пример - сборный род Carabilarva Ponomarenko, предложенный для мезозойских личинок жуков надсемейства Caraboidea, не определимых до семейства (Пономаренко, 1985). Поэтому типовым видом рода является тип надсемейства Carabus granulatus Linnaeus, 1758. Следовательно, Carabilarva является младшим объективным синонимом рода Carabus, но не подлежит синонимизации с ним, пока остается паратаксоном.
Последний пример означает существование еще одной формы множественного подчинения в номенклатуре, помимо уже обсуждавшегося случая, когда разные формы сохранности одного естественного таксона описываются в качестве разных формальных таксонов. Здесь уже один и тот же вид оказывается типовым для множества сборных групп. Действительно, род Carabus одновременно является типовым для семейства Carabidae (жуки-жужелицы), подсемейства Сагаbinае, трибы Carabini и т.д., а для далее неопределимых членов каждого из этих таксонов теоретически могут понадобиться свои сборные группы, да еще разные для личинок и различных изолированных частей тела взрослого жука. Легко представить себе, сколько сборных родов в результате могут оказаться имеющими один и тот же номенклатурный тип. При всей непривычности ситуации, она вряд ли может вызвать серьезные номенклатурные или таксономические проблемы, если мы будем понимать, для чего нужны сборные таксоны. Ведь никому не мешают роды и старшие таксоны неясного положения, почему же отношение к видам неясного положения должно быть иным?
ЗАКЛЮЧЕНИЕ
Еще раз попытаюсь кратко подытожить главное.
Проблемы эволюционного крыла современной биологии, куда я отношу ее разделы, исследующие историю и современное состояние биологического разнообразия, включая их теорию и методологию, на мой взгляд связаны прежде всего с продолжающимися поисками парадигмы. Уже более столетия продолжается спор двух подходов (парадигм) - "элементаристского" и "холистского", но что особенно поразительно, "размежевания" этих двух едва ли совместимых подходов не происходит, а продукты каждого из них сосуществуют в головах и печатной продукции едва ли не большинства ученых, о чем те обычно и не подозревают.
"Элементаристской" парадигме, редуцирующей изучаемую систему к ее элементам, соответствует популяционно-генетическая (синтетическая) теория эволюции, представляющая эволюционный процесс как динамику частот дискретных наследственных детерминант (генов) в популяциях в условиях отбора. Отражением этой динамики и является наблюдаемый процесс эволюции, и ее специфика определяется, во-первых, спецификой генного уровня организации, во-вторых, спецификой отбора. Соответственно эволюционный процесс выступает как существенно равномерный, а единственным регулярным источником дискретности, делающим возможность разбиения продукта эволюции на таксоны, оказывется дивергенция и вымирание. Единственная адекватная этой парадигме таксономическая концепция - кладизм, где таксон возникает в ходе эволюции как продукт дивергенции и вводится нами в систему на том же основании, причем идентификация последовательности дивергенций осуществляется с помощью синапоморфий.
Альтернативная "холистская" парадигма редуцирует эволюционирующую систему к целому и в частности к эпигенотипу, т.е. к системе построения живого организма в онтогенезе. В зеркале эпигенетической теории эволюции сложная, пронизанная переплетающимися взаимосвязями и взаимодействиями живая система предстает напряженным компромиссом противоречивых требований оптимизации разных адаптивных функций. Изменение такой системы оказывается существенно затрудненным и в случае реализации мало предсказуемым и более или менее скачкообразным, а специфика эволюционного процесса определяется прежде всего спецификой эпигенотипа и лишь в существенно меньшей степени - спецификой отбора. Из-за скачкообразности эволюционных изменений биологическое разнообразие, возникающее в ходе такой эволюции, оказывается дискретным, разбитым гиатусами на естественные отдельности, каковые и заслуживают использования в качестве таксонов общей системы. Соответственно, смысл таксономии, адекватной эпигенетической концепции и "холистской" парадигме, состоит в выявлении объективно существующей дискретности и прослеживании гиатусов.
Анализ доступного материала свидетельствует в пользу его соответствия скорее эпигенетической, чем синтетической концепции. Более важной, однако, представляется сама попытка различения концепций и методологий, используемых в анализе эволюционного процесса и его результатов, как восходящих к той или другой из двух конкурирующих парадигм. Представляется, что строгое различение двух парадигм и их следствий позволит не только и даже не столько сделать осмысленный выбор между ними, сколько избавиться от одновременного использования взаимно несовместимых принципов и подходов, т.е. от своего рода плюрализма в одной голове.
Сделанный вывод позволяет несколько продвинуться в выявлении корней и более четком формулировании методологических концепций, используемых на поле эволюционного крыла биологии, и в частности в филогенетике, таксономии и таксономической номенклатуре. Однако вначале нам пришлось рассмотреть некоторые самые общие проблемы научного (и не только) анализа.
Филогенетические выводы, как любое научное исследование, опираются на (1) планирование предстоящего исследования, (2) наблюдение (включая эксперимент), (3) поиск аналогий, (4) создание гипотез о лежащих в основе видимой картины закономерностях и механизмах, (5) попытки фальсификации этих гипотез (преимущественно путем анализа следствий из них), и (6) оценки результатов этих попыток с помощью презумпций, с целью выбора наиболее перспективной гипотезы для дальнейшей работы.
Набор филогенетических презумпций включает презумпцию познаваемости филогенеза и ряд более частных презумпций, которые можно разделить на используемые в анализе групп и в анализе признаков. Анализ групп исследует отношения предков и потомков и опирается прежде всего на палеонтологическую презумпцию для групп. Презумпции, используемые в анализе признаков, далее делятся на две группы соответственно тому, включены ли они в анализ различий или сходств. Анализ различий - это поляризация трансформационных серий, т.е. определение плезиоморфных (исходных) и апоморфных (измененных) состояний признака. Здесь работают, из более важных, палеонтологическая презумпция для признаков, биогенетическая презумпция, презумпции аналогии, необратимости эволюции, функционального совершенства, сложности, рудиментов, и сохраненного распространения признаков. Презумпции анализа сходств используются при решении вопроса, унаследовано ли данное сходство от общего предка или возникло независимо. Сюда относятся прежде всего презумпции парсимонии и взвешенного сходства.
Цель таксономии состоит в создании системы, чьи таксоны осмысленны для максимально широкого круга пользователей и, соответственно, наиболее гомогенны внутри себя и различны между собой. Из трех главных конкурирующих подходов кладизм опирается на синтетическую теорию эволюции, игнорирующую дискретность биоразнообразия. Поэтому кладистическая система учитывает только дивергенции и признает лишь таксоны, охарактеризованные синапоморфиями. Синапоморфия - не признак, а его интерпретация, поэтому кладистическая система не только игнорирует дискретность биоразнообразия, но и излишне гипотетична и привередлива в отборе признаков.
Филистика и фенетика опираются на эпигенетическую теорию эволюции и заняты прослеживанием гиатусов между таксонами. Фенетика использует доступные признаки как таковые и потому слишком близорука, это существенно эмпирический подход. Филистика же занимает промежуточное положение между фенетикой и кладизмом в том, что она пытается отразить полный баланс сходств и различий, включая и те, что еще не изучены. Для этого филистика использует прогностические возможности филогенеза, который выступает в качестве эвристического метода контроля системы, построенной фенетическими методами. Филистика определяет таксон как монофилетический континуум и использует гиатусы для разграничения таксонов, а монофилию как способ оценить, успешно ли было разграничение, или система требует доработки.
В основе таксономической номенклатуры лежат 9 принципов, из которых шесть представляют чистые соглашения для удобства и единообразия в использовании названий таксонов и независимы от используемого таксономического подхода. Остальные три принципа зависят от таксономии. Из них наиболее важен принцип типа, позволяющий стабилизировать названия таксонов, соответствующих понятию монофилетического континуума. Два других таксономически зависимых принципа - принципы иерархии и синонимии, которые могут быть нарушены при работе с дефектным таксономическим материалом, недостаточно охарактеризованным для его полноценной систематизации. В этом случае могут использоваться таксоны неясного положения и паратаксоны разного рода, используемые в рамках специальных систем, параллельных основной системе организмов.
В отличие от принципов синонимии и иерархии, видимо, не существует объективных причин отказываться от соблюдения принципа типа, а тем более от таксономически независимых принципов номенклатуры.
СПИСОК ЛИТЕРАТУРЫ
Алексеев А.С., Дмитриев В.Ю., Пономаренко А.Г. Эволюция таксономического разнообразия // Экосистемные перестройки и эволюция биосферы, вып. 5. М.: Геос, 2001. 126 с.Другие работы А.П. Расницына
English
Главная
Сотрудники
Экспедиции
Ссылки
Публикации
События
Потери науки